Michele PANUCCIO, Nicolantonio AGOSTINI. 2010: Comparison of the water-crossing behavior of Western Marsh Harriers (Circus aeruginosus) and European Honey Buzzards (Pernis apivorus) during autumn migration. Avian Research, 1(1): 30-35. DOI: 10.5122/cbirds.2009.0003
Citation:
Michele PANUCCIO, Nicolantonio AGOSTINI. 2010: Comparison of the water-crossing behavior of Western Marsh Harriers (Circus aeruginosus) and European Honey Buzzards (Pernis apivorus) during autumn migration. Avian Research, 1(1): 30-35. DOI: 10.5122/cbirds.2009.0003
Michele PANUCCIO, Nicolantonio AGOSTINI. 2010: Comparison of the water-crossing behavior of Western Marsh Harriers (Circus aeruginosus) and European Honey Buzzards (Pernis apivorus) during autumn migration. Avian Research, 1(1): 30-35. DOI: 10.5122/cbirds.2009.0003
Citation:
Michele PANUCCIO, Nicolantonio AGOSTINI. 2010: Comparison of the water-crossing behavior of Western Marsh Harriers (Circus aeruginosus) and European Honey Buzzards (Pernis apivorus) during autumn migration. Avian Research, 1(1): 30-35. DOI: 10.5122/cbirds.2009.0003
Comparison of the water-crossing behavior of Western Marsh Harriers (Circus aeruginosus) and European Honey Buzzards (Pernis apivorus) during autumn migration
We investigated the water-crossing behavior of Western Marsh Harriers (Circus aeruginosus), mostly adults, and European Honey Buzzards (Pernis apivorus), mostly juveniles, in relation to wind conditions, time of the day, flocking and age classes, at a watchsite in central Italy during the autumn migration en route to Africa. Although European Honey Buzzards are less suited than Western Marsh Harriers to undertake long sea crossings, they were more inclined to leave the coast when migrating in flocks. Few birds of both species chose to fly along the coast. Western Marsh Harriers, such as European Honey Buzzards migrating alone, undertook the water crossing rather than stopping migration (birds roosting at the site or flying back inland) during the absence of wind and vice versa during head winds. Conversely, European Honey Buzzards migrating in flocks were not affected in their decision (crossing or stopping migration) by wind direction. Both species undertook the water crossing rather than stopping migration during mornings and vice versa during afternoons. Finally, in both species, adults and juveniles showed the same behavior in front of a water barrier. This result was expected in the case of the Western Marsh Harrier but not from the European Honey Buzzard since, in this species, the water-crossing tendency is age dependent with adults avoiding sea crossings. Our study confirms that flocking significantly affects the water-crossing behavior of European Honey Buzzards during migration. Moreover, in this species, inexperience of juveniles and presumably younger adults, about the high energetic costs of long powered flight and about the existence of shorter routes over water, might explain the strong water-crossing tendency shown by migrants independently from their age.
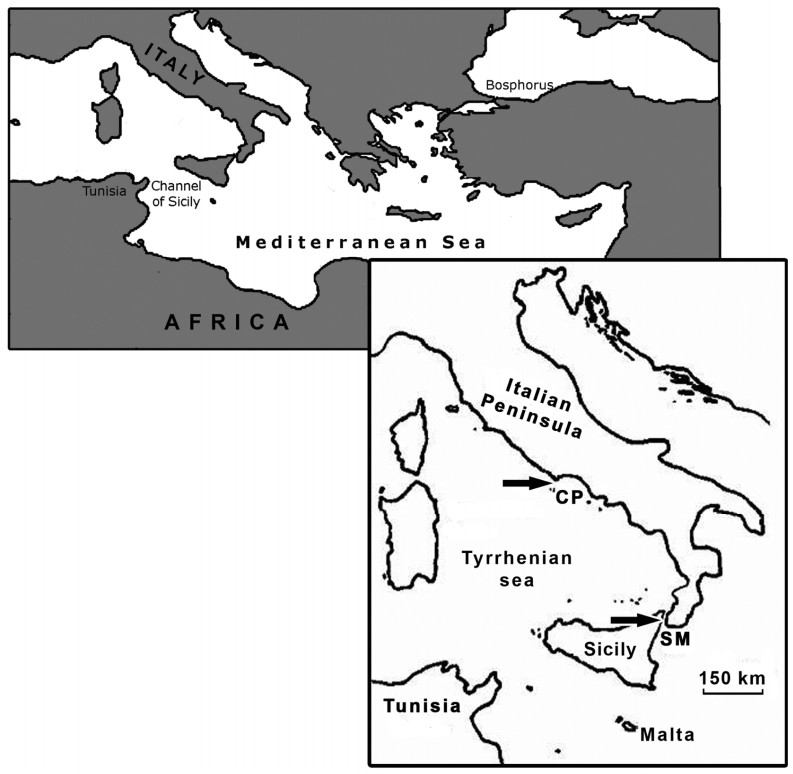
During migration Accipitriformes use mostly soaring flight optimizing the use of thermal currents, avoiding long water crossings to limit powered flight over water in order to reduce energetic costs (Kerlinger, 1989). Several factors influence the decision whether to cross the sea or not and shape the path during the crossing: weather conditions, geography, physiological state of the birds, flocking behavior, time of the day and experience (age dependent), while the risk of mortality probably increases with the absolute distance of the crossing (Kerlinger, 1989; Bildstein, 2006). As well, the flight morphology plays a role; in particular, species with relatively long wings (high aspect ratio = high ratio of wing span squared to wing area; see Kerlinger, 1989) are more suited to undertake crossings of large bodies of water since this feature decreases the induced drag and thus the energy required for powered flight (Kerlinger, 1985). The Western Marsh Harrier (Circus aeruginosus), for instance, is able to cross large bodies of water having high aspect ratio wings; both adults and juveniles migrate on a broad front across the Mediterranean basin (Agostini, 2001; Agostini et al., 2001, 2003; Panuccio et al., 2002). Conversely, in the European Honey Buzzard (Pernis apivorus), a species with a lower wing aspect ratio than the Western Marsh Harrier, the water crossing tendency is age dependent (Agostini and Logozzo, 1995; Agostini et al., 2002; Schmid, 2000; Hake et al., 2003). In particular, during autumn movements through the Central Mediterranean region, thousands of adult (experienced) birds follow the Italian Peninsula avoiding the crossing of the Tyrrhenian Sea (Agostini and Logozzo, 1997; Panuccio et al., 2005). Upon reaching the Strait of Messina between the "toe" of southern Italy and Sicily, they turn west. They will fly across Sicily and then southwest across the Channel of Sicily, the shortest crossing of the Central Mediterranean (approx. 150 km), heading towards Tunisia (Agostini et al., 2000, 2005b; Fig. 1). In contrast, juveniles show a broader front of migration over the sea such as the Western Marsh Harriers. In particular juvenile European Honey Buzzards, migrating later than adults, cannot learn the shortest routes to cross the sea by following experienced birds and tend to move along an innate NE-SW axis between breeding and wintering areas (Agostini and Logozzo, 1995; Agostini et al., 2002; Agostini, 2004). The aim of our study was to compare the water crossing behavior of these two species in relation to wind conditions, the time of the day and the age of migrants, carrying out visual observations at the Circeo Promontory, a watchsite in central Italy. At this site, hundreds of Western Marsh Harriers, mostly adults (Agostini et al., 2001, 2003) and hundreds of European Honey Buzzards, largely juveniles (Corbi et al., 1999; Agostini et al., 2002, 2004), concentrate each autumn.
Figure
1.
Location of study area in the Mediterranean basin (Arrows indicate the Circeo Promontory (CP) and the Strait of Messina (SM))
The Circeo Promontory (41°12′N, 13°03′E) is located at the southernmost point of the Pianura Pontina, reaching 541 m a.s.l. and approximately 500 km northeast of Tunisia (Fig. 1). We used a post (altitude approx. 400 m) in a military zone, on the roof of the ENAV (Ente Nazionale Assistenza al Volo) building; from this post it was possible to observe the flight behavior of birds both inland and over the sea. The Ponziane Islands are nearly always visible from this watch-point; the closer island is Zannone, about 30 km south-southwest of the promontory. A total of 324 hours of observations were carried out between 26 August and 30 September 2002, the peak migration period of these two species, from 09:00 until dusk, aided with binoculars and a telescope. Observations were interrupted only due to rain. As in previous studies investigating the water crossing behavior of these species in relation to the time of the day (Panuccio et al., 2002; Agostini et al., 2005a), each day was divided into three periods (solar time): morning (09:00–11:59), midday (12:00–14:59) and afternoon (15:00–18:00). Meteorological data concerning wind direction were provided by the Meteorological Station of Latina and are available on the website www.ilmeteo.it/dati.htm. The maximum time between the passage of raptors and the meteorological data check was half an hour. Meteorological data were not available for 36 hours. In the analysis we considered crossing and not crossing birds (birds roosting at the site, flying back inland or flying along the coast) using flocks and birds migrating singly as sampling units to evaluate whether the direction of wind and the time of the day affected the choice of these species to undertake the sea crossing or not. Finally, among birds stopping their migration, we did not consider those flying along the coast, but only those roosting at the site and those flying back inland.
Results and discussion
It was possible to follow the movements of 371 flocks (European Honey Buzzard n = 212; Western Marsh Harrier n = 159) and 340 birds migrating singly (European Honey Buzzard n = 180; Western Marsh Harrier n = 160). Only 16 (4.3%) flocks (12 among European Honey Buzzards and 4 among Western Marsh Harriers) and 8 (2.4%) solitary birds (4 European Honey Buzzards and 4 Western Marsh Harriers) left the site flying along the coast. Considering birds that stopped migration (birds roosting at the site or flying back inland) and undertook the crossing of the Tyrrhenian Sea, the behavior of European Honey Buzzards and Western Marsh Harriers did not differ significantly among both flocks (Table 1a; contingency table: χ2 = 1.97, df = 1, p > 0.05) and solitary birds (Table 1b; χ2 = 0.47, df = 1, p > 0.05). During the observation period, tail winds were almost never recorded (11 hours; 3.4%). European Honey Buzzards were not affected by wind conditions when migrating in flocks (Table 2a; contingency table: χ2 = 3.22, df = 2, p > 0.05) while solitary individuals undertook the water crossing rather than stopping migration during the absence of wind and vice versa during head winds (Table 2b; χ2 = 8.15, df = 2, p < 0.05). In contrast, Western Marsh Harriers were affected by wind conditions when migrating both in flocks (Table 2c; χ2 = 7.8, df = 2, p < 0.05) and singly (Table 2d; χ2 = 7.88, df = 2, p < 0.05) with the lowest proportion of birds seen stopping migration during conditions of no wind. Both European Honey Buzzards and Western Marsh Harriers showed the same behavior during all times of the day. In particular, considering both flocks and solitary birds, they undertook the sea crossing rather than stopping migration during the morning. The opposite was true in the afternoon (Table 3a: χ2 = 37.29, df = 2, p < 0.01; Table 3b: χ2 = 19.53, df = 2, p < 0.01; Table 3c: χ2 = 30.65, df = 2, p < 0.01; Table 3d: χ2 = 36.22, df = 2, p < 0.01). It is interesting to note that no wind conditions prevailed during the morning, while the occurrence of head winds was higher at midday and in the afternoon (Table 4; χ2 = 56.94, df = 4, p < 0.001). When comparing the average flock sizes of the two species, the difference was statistically significant between flocks stopping migration (EHB = 4.3 ± 0.43 [SE]; WMH = 7.1 ± 1.5 [SE]; z = 4.67, p < 0.01) but not between those undertaking the crossing (EHB = 5.5 ± 0.45 [SE]; WMH = 4.3 ± 0.37 [SE]). Larger flocks of Western Marsh Harriers formed in the afternoon when they roosted on trees at the site (see also Panuccio and Agostini, 2006). Our results confirm that flocking significantly affects the decision (crossing or not) of European Honey Buzzards when facing a water barrier (Agostini et al., 1994, 2005b) since, unlike flocks, single birds were less inclined to undertake the crossing, hesitating during head winds. Conversely, among Western Marsh Harriers, solitary birds behaved much the same as those flying in flocks. Thus, European Honey Buzzards behaved as reported for raptors that have a higher wing aspect ratio. They were even more inclined than Western Marsh Harriers to undertake the crossing of the water barrier when migrating in flocks. It was possible to determine the age of 1019 birds by observation of their plumage (Forsman, 1999). As reported in a previous study made at this watchsite (Corbi et al., 1999), juveniles (n = 429) outnumbered adults (n = 169) among European Honey Buzzards. Among Western Marsh Harriers, adults (n = 309) outnumbered juveniles (n = 112). When considering the behavior of migrants (crossing; stopping; flying along the coast) in relation to the two age classes, adults behaved like juveniles in both species (Table 5a; contingency table: χ2 = 0.51, df = 2, p > 0.05; Table 5b: χ2 = 3.82, df = 2, p > 0.05). This result was expected in the case of the Western Marsh Harrier since, as mentioned earlier, this is a species which is more adapted to make crossing flights. In contrast, adult European Honey Buzzards, unlike juveniles, were expected to fly along the coast and, as mentioned already, crossing the sea at narrower points (the Strait of Messina and the Channel of Sicily; see also Agostini and Panuccio, 2005) and able to orientate themselves using their navigational abilities and compensating for wind drift (Meyer et al., 2000; Thorup et al., 2003; Agostini et al., 2005b). Our observations suggest that adults passing at the Circeo Promontory each autumn are probably younger, less experienced birds. Similar to the Circeo Promontory, a scarce passage of "adult" European Honey Buzzards was reported over the islands of Malta, Pianosa and Cabrera (Balearic Islands), where mostly juveniles concentrated during autumn migration (Rebassa, 1995; Agostini et al., 2002; Paesani and Politi, 2002). Perhaps, as suggested by Agostini et al.(2002, 2004), juvenile European Honey Buzzards passing over the Circeo Promontory do not have enough experience about the high energetic costs of long powered flight over water and, as younger adults, they do not know the shortest flyway to cross the Central Mediterranean, between western Sicily and Tunisia. Probably for this reason, both juvenile and adult European Honey Buzzards observed in our study behaved similarly, showing a strong tendency to continue migration over the sea. In contrast, our observations do not confirm the hypothesis that the difference in migration strategy between adults and juveniles in this species may partly depend on differences in the timing of migration (Schmid, 2000). In particular, Schmid suggested that since adults mainly migrate across Europe in late August to early September, when it is still possible to travel by soaring flight to a high degree, they use a safer overland flight, since this is compensated by low energetic costs of soaring, compared to flapping flights. Juveniles migrate about two to three weeks later, when meteorological thermal models suggest that conditions for using soaring flights are poorer. Thus, Schmid concluded that they may as well use flapping flights, choosing the shortest way to wintering grounds in West Africa. However, during our study, European Honey Buzzards behaved in the same way during the entire period and, as expected, adults migrated earlier in the season (see also Agostini et al., 2004).
Table
1.
European Honey Buzzards (EHB) and Western Marsh Harriers (WMH) crossing and stopping (birds roosting at the site or flying back inland) migration during the observation period. Birds flying along the coast were not considered in the analysis.
We are grateful to the Aeronautica Militare Italiana for permission to enter a military zone and the ENAV for the permission to use the observation post on the roof of its building. We wish to thank an anonymous referee for his useful comments to an earlier draft of the manuscript.
Agostini N. 2001. Spring migration in relation to sex and age of Marsh Harriers Circusaeruginosus in a central Mediterranean island. Ardeola, 48: 71–73
Agostini N. 2004. Additional observations of age-dependent migration behaviour in western honey-buzzards Pernis apivorus. J Avian Biol, 35: 469–470
Agostini N, Malara G, Neri F, Mollicone D, Melotto S. 1994. Flight strategies of Honey Buzzards during spring migration across the central Mediterranean. Avocetta, 18: 73–76
Agostini N, Logozzo D. 1995. Autumn migration of honey buzzards in southern Italy. J Raptor Res, 29: 275–277
Agostini N, Logozzo D. 1997. Autumn migration of Accipitriformes through Italy en route to Africa. Avocetta, 21: 174–179
Agostini N, Logozzo D, Panuccio M. 2000. The island of Marettimo (Italy), important bird area for the autumn migration of raptors. Avocetta, 24: 95–99
Agostini N, Coleiro C, Corbi F, Di Lieto G, Pinos F, Panuccio M. 2001. Comparative study on the autumn migration of Marsh Harriers at three sites of the central Mediterranean. Vogelwarte, 41: 154–158
Agostini N, Coleiro C, Corbi F, Di Lieto G, Pinos F, Panuccio M. 2002. Water-crossing tendency of juvenile Honey Buzzards during migration. Avocetta, 26: 41–43
Agostini N, Coleiro C, Panuccio M. 2003. Autumn migration of Marsh Harriers across the central Mediterranean in 2002. Ring, 25: 47–52
Agostini N, Coleiro C, Panuccio M. 2004. Analysis of the autumn migration of juvenile honey buzzards (Pernis apivorus) across the central Mediterranean. J Raptor Res, 38: 283–286
Agostini N, Panuccio M. 2005. Analysis of the spatial migration patterns of adult Honey Buzzards (Pernis apivorus) during spring and autumn in the Central Mediterranean. Ring, 27: 29–34
Agostini N, Panuccio M, Massa B. 2005a. Flight behaviour of Honey Buzzards (Pernis apivorus) during spring migration over the sea. Buteo, 14: 3–9
Agostini N, Premuda G, Mellone U, Panuccio M, Logozzo D, Bassi E, Cocchi L. 2005b. Influence of wind and geography on orientation behavior of adult honey buzzards Pernis apivorus during migration over water. Acta Ornithol, 40: 71–74
Bildstein KL. 2006. Migrating Raptors of the World: Their Ecology and Conservation. Cornell University Press, Ithaca, NY
Corbi F, Pinos F, Trotta M, Di Lieto G, Cascianelli D. 1999. La migrazione post-riproduttiva dei rapaci diurni nel promontorio del Circeo (Lazio). Avocetta, 23: 13 (in Italian)
Hake M, Kjellén N, Alerstam T. 2003. Age dependent migration strategy in honey buzzards Pernis apivorus tracked by satellite. Oikos, 103: 385–396
Forsman D. 1999. The raptors of Europe and the Middle East: a Handbook of Field Identification. T & AD Poyser, London
Kerlinger P. 1985. Water-crossing behavior of raptors during migration. Wilson Bull, 97: 109–113
Kerlinger P. 1989. Flight Strategies of Migrating Hawks. University of Chicago Press, Chicago
Meyer KS, Spaar R, Bruderer B. 2000. To cross the sea or to follow the coast? Flight directions and behaviour of migrating raptors approaching the Mediterranean Sea in autumn. Behaviour, 137: 379–399
Paesani G, Politi MP. 2002. Monitoraggio della migrazione autunnale dei rapaci diurni nell'isola di Pianosa (LI). Informamigrans, 10: 6–7 (in Italian)
Panuccio M, Agostini N, Massa B. 2002. Crossing the Tyrrhenian Sea: spring migration of Marsh Harriers (Circus aeruginosus), sex classes and relation to wind conditions. Vogelwarte, 41: 271–275
Panuccio M, Agostini N, Mellone U. 2005. Autumn migration strategies of honey buzzards, black kites, marsh and Montagu's harriers over land and over water in the Central Mediterranean. Avocetta, 29: 27–32
Panuccio M, Agostini N. 2006. Comments on the roosting behaviour of Marsh Harrier during migration. British Birds, 99: 367–368
Rebassa M. 1995. La migraciò postnupcial de rapinyaires diurns a l'Illa de Cabrera: trets geenerals. A.O.B., 10: 11–17 (in Spanish)
Schmid H. 2000. GetrennteWege: Der Herbstzug von juvenilen und adulten Wespenbussarden Pernis apivorus: eine Synthese. Ornith Beob, 97: 191–222 (in German)
Thorup K, Alerstam T, Hake M, Kjellén N. 2003. Bird orientation: compensation for wind drift in migrating raptors is age dependent. Proc R Soc Lond, 270: S8–S11
Table
1.
European Honey Buzzards (EHB) and Western Marsh Harriers (WMH) crossing and stopping (birds roosting at the site or flying back inland) migration during the observation period. Birds flying along the coast were not considered in the analysis.

 DownLoad:
DownLoad:





 Email Alerts
Email Alerts RSS Feeds
RSS Feeds