Roselvy Juárez, Eduardo Chacón-Madrigal, Luis Sandoval. 2020: Urbanization has opposite effects on the territory size of two passerine birds. Avian Research, 11(1): 11. DOI: 10.1186/s40657-020-00198-6
Citation:
Roselvy Juárez, Eduardo Chacón-Madrigal, Luis Sandoval. 2020: Urbanization has opposite effects on the territory size of two passerine birds. Avian Research, 11(1): 11. DOI: 10.1186/s40657-020-00198-6
Roselvy Juárez, Eduardo Chacón-Madrigal, Luis Sandoval. 2020: Urbanization has opposite effects on the territory size of two passerine birds. Avian Research, 11(1): 11. DOI: 10.1186/s40657-020-00198-6
Citation:
Roselvy Juárez, Eduardo Chacón-Madrigal, Luis Sandoval. 2020: Urbanization has opposite effects on the territory size of two passerine birds. Avian Research, 11(1): 11. DOI: 10.1186/s40657-020-00198-6
Urban expansion has been identified as one of the leading drivers of biodiversity change or loss. For birds, urbanization is specifically related to survival, breeding success, and territory size. Understanding how different birds adjust territory size in response to urbanization is essential for their conservation in urban environments and to better understand why some species are lost and others persist under this condition. We evaluated the effect of urbanization on the territory size of an urban avoider species, White-eared Ground-Sparrow (Melozone leucotis), and an urban adapter species, House Wren (Troglodytes aedon), at five Costa Rican sites.
Methods
We measured the size of 30 ground-sparrow and 28 wren territories using a total of 296 h of observation. We followed each individual for at least 1 h per day for at least 2 days of two consecutive years, and geo-referenced their locations. Territory size was estimated using the minimum convex polygon method. We measured the urban surfaces (roads, buildings, any other paved area, soccer fields, lawns, and gardens with short grass) within territories.
Results
Ground-sparrow territories were larger at the highly urbanized site than at the non-urbanized site. Wren territories were larger at the low urbanized site than at the highly urbanized site. We found a positive relationship between urban surface and territory size for the ground-sparrow, but not for the wren.
Conclusions
Our results showed that not all birds adjust territory size in the same way in response to urbanization. We showed that urban avoiders probably need to defend larger territories in urban environments to find all the resources required to survive because urban environments may provide insufficient resources such as food or shelter. Urban adapters on the other hand defend smaller territories in urban environments because even small territories may provide sufficient resources. These results suggest specific behavioral adaptations developed by Neotropical birds inhabiting urban environments.
Territory size and shape are not static over time and may change according to resource abundance and the energy invested in defense (Hixon et al. 1983; Møller 1990; Butchart et al. 1999). Therefore, a balance is expected between territory size and resource abundance or territory size and the time that each individual invests in territorial defense, given that territory size is adjusted according to the benefits obtained from the defense (Brown 1969; Salomonson and Balda 1977; Hixon et al. 1983). For example, in Rufous Hummingbird (Selasphorus rufus) there is a positive correlation between territory size and weight increase, until a threshold over which the increase stops (Carpenter et al. 1983).
In urban environments, some resources may be more abundant than others but of inferior quality (Mennechez and Clergeau 2006). For example, human waste is more abundant in cities, benefiting species that use this resource as part of the diet, resulting in smaller territories inside cities compared to rural or natural habitats (McGowan 2001; Lowry et al. 2013). The opposite is also true, because in urban environments, reduced abundance of natural or specialized resources as specific fruits, flowers, or insects, encourages birds feeding on those resources to defend larger territories to meet their requirements (Mennechez and Clergeau 2006; Weaving et al. 2014). Even generalist species that feed on human waste or human-supplied food may depend on high-quality natural resources to supply all their nutritional and energetic requirements, prompting their necessity to defend larger territories to meet them (Mennechez and Clergeau 2006; Lowry et al. 2013). In general, it is expected that urban avoider species (i.e. species sensitive to human-produced changes in the landscape and with specialized diets) have larger territories inside cities than urban adapter species (i.e. species able to use additional resources provided by humans) (Salomonson and Balda 1977; Marzluff et al. 2016; Ruiz-Sánchez et al. 2017). Considering that not all species respond equally to urbanization and that territory size is not static over time (Møller 1990; MacDougall-Shackleton and Robertson 1995; Holland et al. 2017), we need to understand how different species adjust the size of their territories in response to changes in resource availability generated by humans, to maintain diversity in urban environments.
Our main objective in this study is to test the structural cues hypothesis (Smith and Shugart 1987), which proposes that bird territory size is related to structural habitat features (e.g. urban and natural habitat) as an indicator of resource supply (i.e. food and nesting sites). More specifically, we evaluated two objectives. First, we evaluated if the degree of urbanization is associated with territory size in White-eared Ground-Sparrow (Melozone leucotis) and House Wren (Troglodytes aedon). Second, we evaluated if House Wren territory size varies throughout the year along an urban–rural gradient. We selected White-eared Ground-Sparrow and House Wren as model species because both occur sympatrically along urban–rural gradients in the Central Valley of Costa Rica and both show different affinities for urban environments. White-eared Ground-Sparrow inhabits dense thickets of humid and young secondary forest, coffee plantations, and densely vegetated gardens (Stiles and Skutch 1989; Sandoval and Mennill 2012; Sandoval et al. 2015). Both pair members, male and female, defend the same territory year-round during multiple years (Sandoval et al. 2015), but territory defense appears to be more pronounced during the breeding season when males sing solo songs (Sandoval et al. 2016). Conversely, House Wren is found in open or semi-open areas, from forest edges to urban areas with scattered vegetation (Skutch 1953; Howell and Webb 1995; Johnson 2014). Both male and female House Wren stay in the territory year-round and males sing all year to defend their first territory for life (Skutch 1953; Johnson 2014). Although both species occur throughout the Central Valley of Costa Rica, they are not equally abundant along its urban–rural gradient. White-eared Ground-Sparrow is an urban avoider and achieves moderate densities in natural habitats (Blair 1996; Marzluff and Rodewald 2008). House Wren is an urban adapter and can be common in urban environments with forest patches (Blair 1996; Marzluff and Rodewald 2008).
White-eared Ground-Sparrow and House Wren differ in the benefits obtained by using infrastructure or landscape produced by urbanization. For example, House Wrens forage in the majority of substrates available if these contain invertebrates, and build nests in buildings and other cavities provided by humans (Skutch 1953; Johnson 2014), so they may only need to defend a small space in order to find food and other resources. White-eared Ground-Sparrows, on the other hand, feed mostly in humid leaf litter and require dense thickets to build nests (Sandoval and Mennill 2012), so they may need to defend more space to find food and other resources. Thus, for White-eared Ground-Sparrow, we expected to find larger territories at the urban sites than at the rural ones; meanwhile, for House Wren, we expected to find the opposite pattern. Territory size changes over time according to the status of the breeding cycle; to secure breeding success males need to defend larger territories to protect food resources and mates (Møller 1990; MacDougall-Shackleton and Robertson 1995; Holland et al. 2017). Thus, for House Wren, we expected to find smaller territories during the nonbreeding season than during the breeding season.
Methods
Study area
We conducted this study at five sites within the Central Valley of Costa Rica. (1) Universidad de Costa Rica (9.9378° N, 84.0498° W, Coordinate Reference System: WGS84). The habitat at this location is 1.5 ha forest reserve surrounded by gardens, isolated trees, and buildings. The reserve is a 50-years old secondary growth forest composed of trees such as Ficus spp. (Moraceae), Erythrina poeppigiana (Fabaceae), and Cordia eriostigma (Boraginaceae) with an understory of palms, dense bushes, and some vines (Nishida et al. 2009). Birds at this site are exposed to high human disturbance. (2) Ciudad de la Investigación (9.9390° N, 84.0438° W). The habitat at this location is a mix of a riparian strip, gardens, isolated trees, and buildings. The riparian strip is dominated by trees such as Ficus spp. (Moraceae) and Erythrina poeppigiana (Fabaceae). Birds at this site are exposed to moderate to high human disturbance. (3) Instalaciones Deportivas (9.9452° N, 84.0449° W). The habitat at this location is a mix of small patches of secondary forest mixed with dense thickets of native herbs and exotic grasses, gardens with isolated trees, and some buildings (Biamonte et al. 2011). The secondary forest is dominated by trees such as Ficus spp. (Moraceae), Inga spp., and Cedrela odorata, with an understory of dense bushes and vines. Birds at this site are exposed to moderate human disturbance. (4) Jardín Botánico Lankester (9.8391° N, 83.8903° W). The habitat at this location is a mix of a medium-sized secondary forest, with gardens and a few buildings. The forest is dominated by Ficus spp. (Moraceae) trees with an understory of dense bushes and vines. Birds at this site are exposed to moderate to low human disturbance. (5) Getsemaní de Heredia (10.0329° N, 84.1131° W). The habitat at this location is a mix of a large secondary forest and an abandoned shaded coffee plantation. The forest is dominated by trees such as Ficus spp. (Moraceae), Inga spp. (Fabaceae), Lonchocarpus spp. (Fabaceae), Erythrina spp. (Fabaceae), and Musa spp. plants with an understory of dense bushes and vines. Birds at this site are exposed to very low human disturbance. Hereafter we will refer to these sites as highly urbanized, medium–high urbanized, medium–low urbanized, low urbanized, and non-urbanized sites respectively. These sites were selected because they are exposed to a gradient of anthropogenic disturbance (e.g. human presence, light pollution, and noise) from the highly urbanized to the non-urbanized site, which positively correlates with urban surface (Blair 1996; Warren et al. 2006). We did not sample White-eared Ground-Sparrow at the medium–high urbanized site because only one pair was present in a site of difficult access; nor House Wren at the non-urbanized site because their density was very low and the individuals were hard to follow and observe inside their habitats. For this reason, we excluded these sites from the study for each species.
Fieldwork procedure
At each site, we captured and color-banded at least five males (House Wren) or pairs (White-eared Ground-Sparrow). We followed them during two consecutive breeding seasons. For House Wrens, we only marked males because only they defend territories (Johnson 2014). In total, we followed 32 pairs of White-eared Ground-Sparrows (highly urbanized site = 5, medium–low urbanized site = 6, low urbanized site = 13, and non-urbanized site = 8; 17 pairs were followed both years) and 30 House Wrens (highly urbanized site = 7, medium–high urbanized site = 7, medium–low urbanized site = 6, and low urbanized site = 10; 21 individuals were followed both years). At the beginning of each breeding season (April to July 2015 and 2016), we followed focal individuals on two different days for 1 h per day, between 0500 and 0700 h, when both species were typically more active and easier to observe, for a combined total of 200 h of observation. We also monitored 24 House Wren territories outside the breeding season (from mid-October to the end of November, in 2015 and 2016), on two different days for 1 h per day, between 0500–0700 h, for a combined total of 96 h of observation. While White-eared Ground-Sparrows maintain territories year-round, we studied them only during the breeding season, when their territorial behavior is more pronounced (Sandoval et al. 2015; 2016). Outside the breeding season, pairs are shy and reduce the number of interactions with neighbors, and males do not sing from territory edge perches (Sandoval et al. 2016), rendering the collection of enough data points to estimate territory size difficult. We avoided disturbing focal individuals by following them from a moderate distance (average of 12 m, range: 8–16 m), and by wearing unobtrusive clothing. At the end of every hour of observation, we collected the coordinates of each site used by each focal individual for singing using a Global Positioning System device (GPS, GARMIN model map 62, accuracy = 3 m). Both species sing to defend the territory and the ground-sparrow also uses duets for a similar purpose (Skutch 1953; Johnson 2014; Sandoval et al. 2016). We collected the coordinates only when the GPS precision indicated ≤ 4 m. For White-eared Ground-Sparrow, the number of coordinates collected per territory ranged from 5 to 25 (mean ± SE: 9 ± 0.6); for House Wren, the number of coordinates collected per territory ranged from 5 to 24 (9 ± 0.4).
Urbanization analysis
Because the position of any locality along a landscape gradient not necessarily indicates its degree of urbanization, we corroborated our estimations measuring the proportion of urban surfaces (i.e. roads, buildings, any other paved area, soccer fields, lawns, and gardens with short grass) within a 500 m radius circle around each site sampled. We used as circle center the centroid of all coordinates collected for each territory. Within each circle, we measured the urban surface area by manually drawing polygons over satellite images using the land area calculator in Google Earth Pro. The spatial resolution of the satellite images used was 46 cm, with an eye altitude of 1.30 km. For this analysis, we followed the methods used by Greggor et al. (2016).
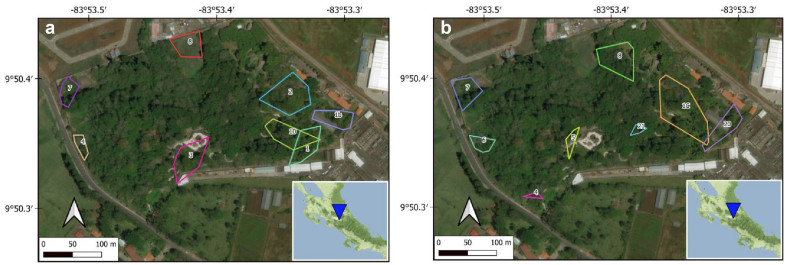
Estimation of territory size
We estimated the territory size of each focal individual (wren) or focal pair (ground-sparrow); using the minimum convex polygon of the GPS coordinates collected during each season per year. This technique of territory estimation has been used to estimate territory sizes in other bird species such as Wilson's Warbler (Cardellina pusilla), Chestnut-crowned Antpitta (Grallaria ruficapilla), and Red-eyed Vireo (Vireo olivaceus; Kattan and Beltran 2002; Marshall and Cooper 2004; Ruiz-Sánchez et al. 2017). We used the "adehabitat" package in R (Calenge 2006), to estimate territory size as the minimum convex polygon using the sets of coordinates of each focal pair. We calculated the minimum convex polygon only for individuals for which at least five sets of coordinates per season (wren) or year (ground-sparrow) were collected (Fig. 1a, b). The average number of coordinates used to estimate the minimum convex polygon is within the minimum required to implement this method to estimate territory size, as well as within the number used in other studies (Marshall and Cooper 2004; Ruiz-Sánchez et al. 2017).
Figure
1.
House Wren and White-eared Ground-Sparrow territories at the low-urbanized site during the breeding season of 2016: a wrens and b ground-sparrows. The inset figure shows the low-urbanized location in Costa Rica
We classified the habitats available in each territory using two categories: (1) urban surface (impervious surface: roads, buildings, and any other paved area; and landscaped surface: lawns, soccer fields, and open gardens with short grass) and (2) natural surface (thickets and secondary forest with well-developed understory). We measured the urban surface areas inside every territory by manually drawing polygons of each type of surface area using satellite images and the land area calculator in Google Earth Pro. Additionally, where necessary, we used field information to distinguish between natural and landscaped surface to draw the polygons. We used satellite images from April 2015 and May 2016, with a spatial resolution of 46 cm, with an eye altitude of 1.30 km.
Statistical analysis
We conducted a linear mixed-effects model to test if the territory size of White-eared Ground-Sparrow varies between the four studied sites (i.e. highly urbanized, medium–low urbanized, low urbanized, and non-urbanized sites). In this analysis, we used the site as the fixed factor and territory size per year per pair as the response variable. We used the pair identity as a random factor to account for the fact that each pair was sampled repeatedly. After the linear mixed-effects model proved that site was a significant factor, we followed up with Tukey's HSD post hoc tests to determine where the differences were. We conducted a linear analysis to test the structural cues hypothesis. We specifically tested if territory size increased with the proportion of urban surface, rather than total urban surface. This analysis corrects for differences in territory size between pairs, allowing us to evaluate if this variable influences territory size. We used parametric tests because we sampled more than 20 territories for this species, and after conducting a square root transformation on territory size, our data follow a normal distribution (Shapiro-Wilks test: W = 0.95, P = 0.12).
We conducted another linear mixed-effects model to test if House Wren territory size varies between season and among the four studied sites (i.e. highly urbanized, medium–high urbanized, medium–low urbanized, and low urbanized sites). In this analysis, we included two fixed factors without interaction: season and site. The response variable was the territory size per season per individual. We used the individual identity as a random factor to account for the fact that each male was sampled repeatedly. Likewise, after the linear mixed-effects model showed that site was a significant factor, we used Tukey's HSD post hoc tests to determine differences between sites. Similar to the model for the White-eared Ground-Sparrow, we then conducted a linear regression to test the structural cues hypothesis. We used parametric tests because tests showed normality for House Wren data (Shapiro-Wilks test: W = 0.96, P = 0.40). In both linear analyses, we used as the Y variable the average of the territory size per pair (White-eared Ground-Sparrow) or per individual (House Wren). We used the R language and environment 3.3.3 for all statistical analyses (R Core Team 2017). We report average ± SE for all our measurements.
Results
Based on our 500 m radius circle, our urbanization analysis supports the classification of the study sites from more to less urban sites: urban surface comprised 66% of the highly urbanized site, 57% of the medium–high urbanized site, 52% of the medium–low urbanized site, 22% of the low urbanized site, and only 1% of the non-urbanized site. We estimated territory size for 30 White-eared Ground-Sparrow territories and 28 House Wren territories for both years combined. These data differ from the number of territories monitored (32 and 30 respectively) because for some focal individuals we were unable to collect a minimum of five sets of coordinates per season (wren) or year (ground-sparrow), preventing us to estimate territory size. White-eared Ground-Sparrow territory size was 0.18 ± 0.03 ha, n = 30. House Wren territory size was 1.8 times larger during the breeding season (0.11 ± 0.01 ha, n = 27) than during the non-breeding (0.06 ± 0.01 ha, n = 24; F1, 55 = 17.24, P < 0.001).
Effect of urbanization on territory size
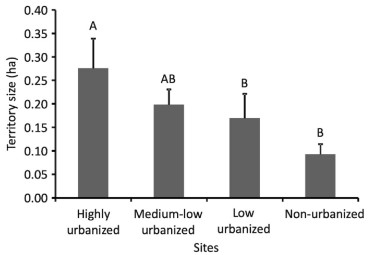
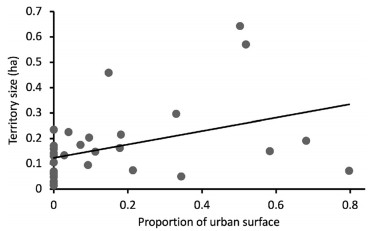
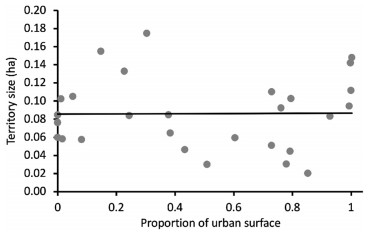
White-eared Ground-Sparrow territory size varied according to the degree of urbanization (F3, 27 = 2.38, P = 0.02, Fig. 2). Territories were 3.1 times larger at the highly urbanized site (0.28 ± 0.04 ha, n = 5) in comparison to the non-urbanized site (0.09 ± 0.02 ha, n = 6; post hoc test: P = 0.01, Fig. 2). We also found this pattern between the highly urbanized site and the low urbanized site, where territories were 1.8 times smaller (0.09 ± 0.01 ha, n = 6; post hoc test: P = 0.03, Fig. 2). For all other pair comparisons between territory sizes according to the degree of urbanization per studied site, we did not find any difference (post hoc test: P > 0.06 for all comparisons, Fig. 2). White-eared Ground-Sparrow territory size increased as the proportion of urban surface inside the territory increased (r2 = 0.15, P = 0.03, Fig. 3).
Figure
2.
Territory size of White-eared Ground-Sparrow at different sites in the Central Valley of Costa Rica. Error bars are standard errors around the mean. The capital letter above the bars indicates statistical differences between those sites. Sample size per site, from left to right, is 5, 6, 13, and 7 respectively
Figure
3.
Relationship between the proportion of urban surface and the territory size of White-eared Ground-Sparrow. Measurements were collected at different sites in the Central Valley of Costa Rica during 2015 and 2016 (n = 30)
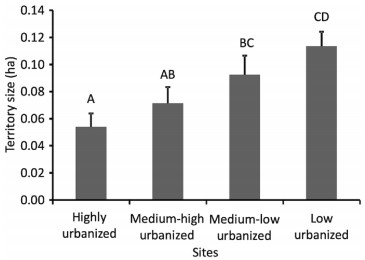
House Wren territory size varied according to the degree of urbanization (F3, 24 = 5.75, P = 0.004, Fig. 4). We found 2.2 times smaller territories at the highly urbanized site in comparison to the low urbanized site (0.05 ± 0.01 ha, n = 7; 0.12 ± 0.01 ha, n = 9; post hoc test: P = 0.001, Fig. 4). We also found this pattern between the highly urbanized site and the medium–low urbanized site, where territories were 1.8 times smaller (0.09 ± 0.01 ha, n = 6; post hoc test: P = 0.03, Fig. 4) and between the medium–high urbanized site and the low urbanized site, where territories were 1.6 times smaller (0.07 ± 0.01 ha, n = 6; post hoc test: P = 0.008, Fig. 4). For all other pair comparisons between territory sizes according to the degree of urbanization per studied site, we did not find any difference (post hoc test: P > 0.06 for all comparisons, Fig. 4). House Wren territory size was not related to the proportion of urban surface inside the territory (r2 = 0.00, P = 0.91, Fig. 5).
Figure
4.
Territory size of House Wren at different sites in the Central Valley of Costa Rica. Error bars are standard errors around the mean. The capital letter above the bars indicates statistical differences between those sites. Sample size per site, from left to right, is 7, 6, 6, and 9 respectively
Figure
5.
Relationship between the proportion of urban surface and the territory size of House Wren. Measurements were collected at different sites in the Central Valley of Costa Rica during 2015 and 2016 (n = 28)
Variation of territory size among birds due to urbanization
We found clear differences in the variation of territory size according to the degree of urbanization in the two studied species. White-eared Ground-Sparrow had larger territories and House Wren had smaller territories at the highly urbanized site. Feeding habits may be the factor driving this pattern. For certain species like some raptors and waterbirds, urban environments provide more food (Frixione et al. 2012; Boggie and Mannan 2014), while for others that require some specialized food at least in some part of their life, like European Starling (Sturnus vulgaris), urban environments may not provide all necessary dietary components (McGowan 2001; Mennechez and Clergeau 2006). Flexibility in feeding habits may allow urban adapters such as House Wren to have smaller territories in urban environments because they can use multiple food and habitat resources (Skutch 1953; Lowry et al. 2013). Meanwhile, a species with more limited feeding habits (e.g. an obligate ground feeder such as White-eared Ground-Sparrow) may need larger territories to obtain all the required food. Although territory size is related to body size, with larger birds defending larger territories (Price 1984; Weaving et al. 2014), a pattern also confirmed in this study, this cannot explain the opposite patterns that we found for our two focal species along an urban gradient.
Additionally, resource availability (e.g. nesting places) may be another factor driving the opposite pattern of the effects of urbanization on the territory size of ground-sparrows and wrens. For secondary cavity-nesting species like House Wren (Skutch 1953; Johnson 2014), urban environments may increase the availability of places for nesting since buildings and other human structures provide many holes to nest (Skutch 1953; Emlen 1974). Conversely, for species that nest on the ground and in dense thickets like White-eared Ground-Sparrow (Sandoval and Mennill 2012), urban environments may offer fewer nesting places (Emlen 1974; Mills et al. 1989; Vignoli et al. 2013). The positive relationship we found between ground-sparrow territory size and the proportion of urban surface provides additional support for the relevance of resource availability in determining territory size (Eberhard and Ewald 1994; Marshall and Cooper 2004; Weaving et al. 2014; Ruiz-Sánchez et al. 2017), because individuals inhabiting highly urbanized territories may need to protect larger areas in order to secure all necessary resources to survive and reproduce.
Density is another factor that may determinate territory size for both species (Brown 1969; Boggie and Mannan 2014; Weaving et al. 2014). It is expected that an urban avoider like White-eared Ground-Sparrow achieves its highest abundances in more natural environments (Blair 1996; Marzluff and Rodewald 2008). Therefore, at urban sites, where ground-sparrows apparently are less abundant (Biamonte et al. 2011), they have more area available, allowing them to defend larger territories. An urban adapter like House Wren, on the other hand, achieves its highest abundance in suburban and urban environments with some forest patches (Skutch 1953; Blair 1996; Boggie and Mannan 2014), where they defend smaller territories compared to natural sites (McGowan 2001; Johnson 2014). The fact that in urban environments ground-sparrows had the largest territories while wrens had the smallest territories compared to the low-urbanized environments provides support for the influence of population abundance in the determination of territory size in these two passerines. However, it is also plausible that the difference in territory size between urban and low-urbanized environments is driving the difference of densities between those environments, as has been suggested to explain differences in densities of several species of antpittas (Kattan and Beltran 2002).
Variation of House Wren territory size across seasons
House Wren territories in the Costa Rican Central Valley were larger during the breeding season than outside the breeding season (approximately 83% larger). In line with our results, larger territories during the breeding season have been reported in several passerine species like Yellowhammer (Emberiza citronella), Barn Swallow (Hirundo rustica), and Wilson's Warbler (e.g. Møller 1990; Ruiz-Sánchez et al. 2017) and non-passerine species like Little Owl (Athene noctua) and Common Cuckoo (Cuculus canorus; e.g. Finck 1990; Williams et al. 2016). For wrens, similar results have been reported in Pacific Wren (Troglodytes pacificus; Hejl et al. 2002) and Marsh Wren (Cistothorus palustris; Kroodsma and Verner 2013). Larger territories during the breeding season, especially at the beginning when females are more fertile, may help males to reduce the risk of the female being fertilized by another male, and therefore lose reproduction opportunities (Møller 1990, 1992). In polygynous species like House Wren, large territories during the breeding season also may allow males more mating opportunities because this may increase the probability of more females settling to breed (Dunn 1992; Johnson 2014; Alcock 2016). Additionally, larger territories may be necessary to access the food resources that a breeding pair requires (Salomonson and Balda 1977; Ruiz-Sánchez et al. 2017). This, in turn, reduces the time that nests are left unattended, thus decreasing nest predation (Beissinger et al. 1998), or infanticide, as has been reported in unattended House Wren nests (Freed 1986; Kermott et al. 1991; Kattan 2016). Larger territories also reduce the need to enter another pair's territory to obtain food, and this reduces territorial conflicts that may result in physical damage to the parents.
Conclusions
Our results regarding the effects of urbanization on the territory size of White-eared Ground-Sparrow and House Wren showed that urbanization has opposite effects on the territory size of these two passerines with different responses to urban development: White-eared Ground-Sparrow has larger territories in urban environments while House Wren shows the opposite pattern. Our results of the annual variation in House Wren territory size support the defense of larger territories during the breeding season, and this may help House Wren males to secure the highest reproductive success. Larger territories may ensure mate fidelity and also prevent the premature death of the pair's offspring by increasing nest and juvenile protection. These results are important for urban management practices because they provide evidence about the habitat structure (proportion of urban and natural surface) and size necessary for the conservation of species with different degree of tolerance to urban development. Our results also highlight the relevance of dense thicket, a habitat generally underappreciated, but important for the conservation of urban adapter and avoider species alike (Sandoval et al. 2019). For an urban avoider species such as White-eared Ground-Sparrow whose natural habitat includes dense understories (Sandoval and Mennill 2012; Sandoval et al. 2016), dense thickets inside urban areas constitute an irreplaceable trophic substratum for roosting, breeding and feeding. This habitat also provides resources for other species of birds, mammals, and arthropods specialized or restricted to this habitat inside urban areas to survive (Sandoval et al. 2019).
Acknowledgements
We are grateful to C. Méndez, M. Villarreal, and O. Sandoval for field assistance. We thank M. Fernández, Eben Goodale, and two anonymous reviewers for helpful comments on early drafts of the manuscript. We thank J. van Dort for useful comments on a previous version of the manuscript and for improving English writing and grammar.
Authors' contributions
LS secured funding for field work. RJ and LS conceived the idea and conzducted the research. RJ, LS, and ECM analyzed the data, and wrote and edited the paper. All authors read and approved the final manuscript.
Funding
RJ gratefully acknowledges a grant in the form of the scholarship Julio E. Sánchez para el Avance de la Ornitología from Unión de Ornitólogos de Costa Rica. LS would like to thank Escuela de Biología, Universidad de Costa Rica, for the partial time support, and to Vicerrectoría de Investigación for the support under the project number B5241 Ecología de la comunicación acústica en aves de ambientes urbanos. None of our funders had any input into the content of the manuscript, nor did they require the approval of the manuscript before submission.
Availability of data and materials
The datasets used and/or analyzed during the current study are available from the corresponding author on reasonable request.
Ethics approval and consent to participate
This study was approved by Vicerrectoria de Investigación, Universidad de Costa Rica number N-111-B5-241. Birds were handled only by two of the authors, both of whom have demonstrable experience banding birds.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Alcock J. Avian mating and social behavior. In: Lovette IJ, Fitzpatrick JW, editors. The Cornell Lab of Ornithology Handbook of bird biology. Hoboken: Wiley Press; 2016. p. 313–53.
Beissinger SR, Tygielski S, Elderd B. Social constraints on the onset of incubation in a neotropical parrot: a nestbox addition experiment. Anim Behav. 1998;55:21–32.
Biamonte E, Sandoval L, Chacón E, Barrantes G. Effect of urbanization on the avifauna in a tropical metropolitan area. Landsc Ecol. 2011;26:183–94.
Boggie MA, Mannan RW. Examining seasonal patterns of space use to gauge how an accipiter responds to urbanization. Landsc Urban Plan. 2014;124:34–42.
Blair RB. Land use and avian species diversity along an urban gradient. Ecol Appl. 1996;6:506–19.
Brown JL. Social organization and behavior of the Mexican Jay. Condor. 1963;65:126–53.
Brown JL. Territorial behavior and population regulation in birds: a review and re-evaluation. Wilson Bull. 1969;81:293–329.
Butchart SH, Seddon N, Ekstrom JM. Polyandry and competition for territories in bronze-winged jacanas. J Anim Ecol. 1999;68:928–39.
Carpenter FL, Paton DC, Hixon MA. Weight gain and adjustment of feeding territory size in migrant hummingbirds. Proc Natl Acad Sci. 1983;80:7259–63.
Calenge C. The package "adehabitat" for the R software: a tool for the analysis of space and habitat use by animals. Ecol modell. 2006;197:516–9.
Duca C, Marini MÂ. Territorial system and adult dispersal in a cooperative-breeding tanager. Auk. 2014;131:32–40.
Dunn PO. Do male birds adjust territory size to the risk of cuckoldry? Anim Behav. 1992;43:857–9.
Eberhard JR, Ewald PW. Food availability, intrusion pressure and territory size: an experimental study of Anna's hummingbirds (Calypte anna). Behav Ecol Sociobiol. 1994;34:11–8.
Emlen JT. An urban bird community in Tucson, Arizona: derivation, structure, regulation. Condor. 1974;76:184–97.
Finck P. Seasonal variation of territory size with the little owl (Athene noctua). Oecologia. 1990;83:68–75.
Freed LA. Territory takeover and sexually selected infanticide in tropical house wrens. Behav Ecol Sociobiol. 1986;19:197–206.
Frixione MG, Casaux R, Villanueva C, Alarcón PA. A recently established Kelp Gull colony in a freshwater environment supported by an inland refuse dump in Patagonia. Emu. 2012;112:174–8.
Greggor AL, Clayton NS, Fulford AJ, Thornton A. Street smart: faster approach towards litter in urban areas by highly neophobic corvids and less fearful birds. Anim Behav. 2016;117:123–33.
Halkin SL, Linville SU. Northern Cardinal Cardinalis cardinalis. In: Poole AF, Gill FB, editors. Birds of North America, version 2.0. Ithaca: Cornell Lab of Ornithology; 1999. .
Hejl SJ, Holmes JA, Kroodsma DE. Winter Wren Troglodytes hiemalis. In: Poole AF, Gill FB, editors. Birds of North America, version 2.0. Ithaca: Cornell Lab of Ornithology; 2002. .
Hixon MA, Carpenter FL, Paton DC. Territory area, flower density, and time budgeting in hummingbirds: an experimental and theoretical analysis. Am Nat. 1983;122:366–91.
Holland AE, Byrne ME, Bryan AL, DeVault TL, Rhodes OE, Beasley JC. Fine-scale assessment of home ranges and activity patterns for resident black vultures (Coragyps atratus) and turkey vultures (Cathartes aura). PLoS ONE. 2017;12:e0179819.
Howell SNG, Webb S. A guide to the birds of Mexico and northern Central America. New York: Oxford University Press; 1995.
Johnson LS. House Wren Troglodytes aedon. In: Poole AF, editor. Birds of North America, version 2.0. Ithaca: Cornell Lab of Ornithology; 2014. .
Kattan GH. Heterospecific infanticidal behavior by Southern House Wrens (Troglodytes aedon musculus) suggests nest site competition. Wilson J Ornithol. 2016;128:899–903.
Kattan GH, Beltran JW. Rarity in antpittas: territory size and population density of five Grallaria spp in a regenerating habitat mosaic in the Andes of Colombia. Bird Conserv Int. 2002;12:231–40.
Kermott LH, Johnson LS, Merkle MS. Experimental evidence for the function of mate replacement and infanticide by males in a north-temperate population of House Wrens. Condor. 1991;93:630–6.
Kraaijeveld K, Dickinson JL. Family-based winter territoriality in western bluebirds, Sialia mexicana: the structure and dynamics of winter groups. Anim Behav. 2001;61:109–17.
Kroodsma DE, Verner J. Marsh Wren Cistothorus palustris. In: Poole AF, editor. Birds of North America, version 2.0. Ithaca: Cornell Lab of Ornithology; 2013. .
Lowry H, Lill A, Wong B. Behavioural responses of wildlife to urban environments. Biol Rev. 2013;88:537–49.
MacDougall-Shackleton EA, Robertson RJ. Mate guarding tactics used by great crested flycatchers. Wilson Bull. 1995;107:757–61.
Marshall MR, Cooper RJ. Territory size of a migratory songbird in response to caterpillar density and foliage structure. Ecology. 2004;85:432–45.
Marzluff JM, Rodewald A. Conserving biodiversity in urbanizing areas: nontraditional views from a bird's perspective. Cit Environ. 2008;1:1–27.
Marzluff JM, DeLap JH, Oleyar MD, Whittaker KA, Gardner B. Breeding dispersal by birds in a dynamic urban ecosystem. PLoS ONE. 2016;11:e0167829.
McGowan KJ. Demographic and behavioral comparisons of suburban and rural American Crows. In: Marzluff JM, Bowman R, Donelly R, editors. Avian ecology and conservation in an urbanizing world. Norwell: Kluwer Academic Press; 2001. p. 365–81.
Mennechez G, Clergeau P. Effect of urbanisation on habitat generalists: starlings not so flexible? Acta Oecol. 2006;30:182–91.
Mills GS, Dunning JB, Bates JM Jr. Effects of urbanization on breeding bird community structure in southwestern desert habitats. Condor. 1989;91:416–28.
Møller AP. Changes in the size of avian breeding territories in relation to the nesting cycle. Anim Behav. 1990;40:1070–9.
Møller AP. Relative size of avian breeding territories and the risk of cuckoldry. Anim Behav. 1992;43:860–1.
Nice MM. The role of territory in bird life. Am Midl Nat. 1941;26:441–87.
Nishida K, Nakamura I, Morales C. Plants and butterflies of a small urban preserve in the Central Valley of Costa Rica. Rev Biol Trop. 2009;57:31–67.
Osborne DR, Bourne GR. Breeding behavior and food habits of the Wattled Jacana. Condor. 1977;79:98–105.
Price TD. Sexual selection on body size, territory and plumage variables in a population of Darwin's finches. Evolution. 1984;38:327–41.
R Core Team. R: A language and environment for statistical computing. Vienna: R Foundation for Statistical Computing; 2017. .
Ruiz-Sánchez A, Renton K, Rueda-Hernández R. Winter habitat disturbance influences density and territory size of a Neotropical migratory warbler. J Ornithol. 2017;158:63–73.
Salomonson MG, Balda RP. Winter territoriality of Townsend's Solitaires (Myadestes townsendi) in a pinon-juniper-ponderosa pine ecotone. Condor. 1977;79:148–61.
Sandoval L, Mennill DJ. Breeding biology of White-eared Ground-sparrow (Melozone leucotis), with a description of a new nest type. Ornitol Neotrop. 2012;23:225–34.
Sandoval L, Dabelsteen T, Mennill DJ. Transmission characteristics of solo songs and duets in a neotropical thicket habitat specialist bird. Bioacoustics. 2015;24:289–306.
Sandoval L, Méndez C, Mennill DJ. Vocal behaviour of White-eared Ground-sparrows (Melozone leucotis) during the breeding season: repertoires, diel variation, behavioural contexts, and individual distinctiveness. J Ornithol. 2016;157:1–12.
Sandoval L, Morales CO, Ramírez-Fernández JD, Hanson P, Murillo-Hiller R, Barrantes G. The forgotten habitats in conservation: early successional vegetation. Rev Biol Trop. 2019;67:S36–52.
Skutch AF. Life history of the southern house wren. Condor. 1953;55:121–49.
Smith TM, Shugart HH. Territory size variation in the ovenbird: the role of habitat structure. Ecology. 1987;68:695–704.
Stiles FG, Skutch AF. A guide to the birds of Costa Rica. Ithaca: Cornell University Press; 1989.
Tinbergen N. The function of sexual fighting in birds; and the problem of the origin of "territory". Bird Band. 1936;7:1–8.
Vignoli L, Scirè S, Bologna MA. Rural–urban gradient and land use in a millenary metropolis: how urbanization affects avian functional groups and the role of old villas in bird assemblage patterning. Web Ecol. 2013;13:49–67.
Warren PS, Katti M, Ermann M, Brazel A. Urban bioacoustics: it's not just noise. Anim Behav. 2006;71:491–502.
Weaving MJ, White JG, Hower K, Isaac B, Cooke R. Sex-biased space-use response to urbanization in an endemic urban adapter. Landsc Urban Plan. 2014;130:73–80.
Williams HM, Willemoes M, Klaassen RH, Strandberg R, Thorup K. Common Cuckoo home ranges are larger in the breeding season than in the non-breeding season and in regions of sparse forest cover. J Ornithol. 2016;157:461–9.
Woltmann S, Terrill RS, Miller MJ, Brady ML. Chestnut-backed Antbird Poliocrania exsul. In: Schulenberg TS, editor. Neotropical Birds Online, version 1.0. Ithaca: Cornell Lab of Ornithology; 2010. .
Woltmann S, Sherry TW. High apparent annual survival and stable territory dynamics of Chestnut-backed Antbird (Myrmeciza exsul) in a large Costa Rican rain forest preserve. Wilson J Ornithol. 2011;123:15–23.
Berlusconi, A., Castiglione, G., Clerici, E. et al. Heterospecific territorial defense in tit species varies according to breeding habitat overlap. Behavioral Ecology, 2025, 36(4): araf082.
DOI:10.1093/beheco/araf082
2.
Cueva, L., Fuchs, E.J., Barrantes, G. et al. Effect of Spatial and Temporal Urban Isolation on the Genetic Diversity, Acoustic Variation, and Morphological Characteristics of an Urban Survivor Bird Species. Ecology and Evolution, 2025, 15(4): e70972.
DOI:10.1002/ece3.70972
3.
Menon, M.. Urban Birds and Adaptive Behaviours. Animal Behavior in the Tropics Vertebrates, 2025.
DOI:10.1007/978-981-97-7324-4_17
4.
Dayananda, S.K., Athukorala, H.F., Peabotuwage, I. et al. Rubber plantations are impermeable to an avian understory specialist in Sri Lanka. Movement Ecology, 2024, 12(1): 47.
DOI:10.1186/s40462-024-00484-8
5.
Sheng, Y., Lu, M., Bai, J. et al. Ecological drivers of nesting behavior in a subtropical city: An observational study on spotted doves. Ecology and Evolution, 2024, 14(7): e11655.
DOI:10.1002/ece3.11655
6.
Villarreal, M., Barrantes, G., Sandoval, L. SONG FREQUENCY SHIFTS IN AN URBAN BIRD SPECIES OPTIMIZE ACOUSTIC TRANSMISSION INSIDE NOISY URBAN AREAS | [Los cambios en la frecuencia del canto en una especie de ave urbana optimizan la transmisión acústica en áreas urbanas ruidosas]. Ornitologia Neotropical, 2024, 35(2): 80-86.
DOI:10.58843/ornneo.v35i2.1363
7.
Cueva, L., Bartolo, R., Sandoval, L. The Acoustic adaptation hypothesis does not support the occurrence of common songs in a neotropical urban bird species. Bioacoustics, 2024, 33(6): 510-526.
DOI:10.1080/09524622.2024.2422091
8.
Vargová, V., Balogová, M., Pristašová, P. et al. Spatiotemporal dynamics in the roosting ecology of the green toad: Implications for urban planning and nature conservation. Journal for Nature Conservation, 2024.
DOI:10.1016/j.jnc.2023.126543
9.
Beck, M.L., Sewall, K.B., Akҫay, Ҫ. Experimental manipulation of chest spotting alters territorial aggression in urban and rural song sparrows. Behavioral Ecology and Sociobiology, 2023, 77(12): 136.
DOI:10.1007/s00265-023-03396-6
10.
Simberloff, R.A., Phillips, J.N., Derryberry, G.E. et al. Communication distance predicts territory size: implications for an urban songbird. Animal Behaviour, 2023.
DOI:10.1016/j.anbehav.2023.05.016
11.
Khan, I.A., Kumar, A., Bhatt, D. et al. Tree cover and built-up area regulate the territory size in Eurasian Magpie Pica pica in Ladakh, India. Journal of Threatened Taxa, 2023, 15(7): 23521-23528.
DOI:10.11609/jott.8214.15.7.23521-23528
12.
Lane, S.J., VanDiest, I.J., Brewer, V.N. et al. Indirect effects of urbanization: consequences of increased aggression in an urban male songbird for mates and offspring. Frontiers in Ecology and Evolution, 2023.
DOI:10.3389/fevo.2023.1234562
13.
Rodríguez-Bardía, M., Fuchs, E.J., Barrantes, G. et al. Genetic structure in neotropical birds with different tolerance to urbanization. Scientific Reports, 2022, 12(1): 6054.
DOI:10.1038/s41598-022-09961-9
14.
Hope, S.F., Hopkins, W.A., Angelier, F. Parenting in the city: effects of urbanization on incubation behaviour and egg temperature in great tits, Parus major. Animal Behaviour, 2022.
DOI:10.1016/j.anbehav.2022.09.004
15.
Amorim, P.S., Guaraldo, A.C., Diniz, P. Horneros consider their neighbors as precious foes regardless of territory size and human disturbance. Behavioural Processes, 2022.
DOI:10.1016/j.beproc.2023.104942
16.
Lane, S.J., Sewall, K.B. What about Females? Urban Female Song Sparrows Elevate Aggressive Signaling Compared to Rural. Integrative and Comparative Biology, 2022, 62(3): 487-495.
DOI:10.1093/icb/icac106
17.
Goodchild, C.G., VanDiest, I., Lane, S.J. et al. Variation in Hematological Indices, Oxidative Stress, and Immune Function Among Male Song Sparrows From Rural and Low-Density Urban Habitats. Frontiers in Ecology and Evolution, 2022.
DOI:10.3389/fevo.2022.817864
18.
Juárez, R., Ruiz-Gutiérrez, V., Sandoval, L. Surviving in cities: The case of a year-round territorial bird in the Neotropics. Journal of Urban Ecology, 2022, 8(1): juac006.
DOI:10.1093/jue/juac006
19.
Hamzaj, T., Graham, B., Bonilla, K. et al. Intruder familiarity and not duet similarity influences the territory defense in a year-round territorial bird species. Avian Research, 2022.
DOI:10.1016/j.avrs.2022.100032
20.
Chen, X., Zhang, Q., Lan, S. et al. Nest predation pressure on Chinese Bulbuls decreases along the urbanization gradient in Hangzhou, China. Avian Research, 2022.
DOI:10.1016/j.avrs.2022.100049
21.
Juárez, R., Angulo Irola, M.P., Carman, E.M. et al. Territory size, population density, and natural history of Cabanis’s Ground Sparrow, an endemic species found in urban areas. Ornithology Research, 2021, 29(4): 227-239.
DOI:10.1007/s43388-021-00076-9
22.
Wang, L., Nabi, G., Yin, L. et al. Birds and plastic pollution: recent advances. Avian Research, 2021, 12(1): 59.
DOI:10.1186/s40657-021-00293-2
23.
Diniz, P., Duca, C. Anthropogenic noise, song, and territorial aggression in southern house wrens. Journal of Avian Biology, 2021, 52(10): e02846.
DOI:10.1111/jav.02846
24.
Brunner, N., Kühleitner, M., Renner-Martin, K. Bertalanffy-Pütter models for avian growth. Plos One, 2021, 16(4 April): e0250515.
DOI:10.1371/journal.pone.0250515
25.
Méndez, C., Barrantes, G., Sandoval, L. The effect of noise variation over time and between populations on the fine spectrotemporal characteristics of different vocalization types. Behavioural Processes, 2021.
DOI:10.1016/j.beproc.2020.104282
Other cited types(0)
Catalog
Luis Sandoval
1.
Escuela de Biología, Universidad de Costa Rica, CP: 11501-2060, San Pedro, Montes de Oca, San José, Costa Rica

 DownLoad:
DownLoad:





 Email Alerts
Email Alerts RSS Feeds
RSS Feeds