Anthropogenic nesting materials and reproductive performance of Chinese Bulbuls (Pycnonotus sinensis) along the urbanization gradient in Hangzhou, China
Urbanization induced by human activities presents both challenges and adaptive opportunities for wildlife. One notable impact of urban sprawl is the vast amount of waste it produces, which has discernible effects on wildlife. Interestingly, an emerging trend has been observed that birds are incorporating anthropogenic materials into their nests. However, the relationship between anthropogenic nesting materials (ANMs) and the reproductive performance of urban birds is unclear. In this study, we investigated the relationship between the incorporation of ANMs into nests and the reproductive performance of Chinese Bulbuls (Pycnonotus sinensis) by monitoring and collecting data on 136 breeding nests in Hangzhou, China. We gathered data on seven reproductive traits of Chinese Bulbuls, including laying date, incubation period, nestling period, clutch size, egg volume, hatching success rate, and fledging success rate. We then calculated the urbanization synthetic index as a measure of the level of urbanization and examined its relationship with the proportion and weight of ANMs. Through examination of nest components, we observed significant increase in the proportion and weight of ANMs with the urbanization synthetic index. Notably, we found a higher hatching success rate of Chinese Bulbuls with an increasing proportion of ANMs. However, the inclusion of ANMs in nests was not correlated with other reproductive traits of Chinese Bulbuls. Overall, the use of ANMs by Chinese Bulbuls is consistent with the adaptive hypothesis and the availability hypothesis. Further studies should use controlled experiments to investigate the impact of ANMs on avian reproductive success.
Nests are crucial in determining the reproductive success of birds (Hansell, 2005; Collias and Collias, 2016). The nest protects the eggs and offspring from predators and environmental stressors (Deeming and Reynolds, 2016). Urban birds have been found to integrate anthropogenic nesting materials (ANMs) into their nests to adapt to the city environment (Wang et al., 2009; Reynolds et al., 2019; Jagiello et al., 2022). However, such new ANMs can yield mixed consequences for birds. For instance, the incorporation of ANMs affects the cooling dynamics of Clay-colored Thrush (Turdus grayi) nests (Corrales-Moya et al., 2021), but can simultaneously decrease nest survival rates (Corrales-Moya et al., 2023). Some ANMs like cigarette butts might deter parasites, affect the physical well-being of adult birds, but elevate the reproductive costs in urban settings (Suárez-Rodríguez et al., 2013, 2017). Jagiello et al. (2022) also showed that the higher the ANMs' proportion in the Blue Tits' (Cyanistes caeruleus) nests in urban habitats, the lower the number of hatchlings.
Several hypotheses have been proposed to explain why birds use ANMs to build nests (Reynolds et al., 2019; Jagiello et al., 2022, Jagiello et al., 2023). First, the availability hypothesis holds that urban birds are left with little choice due to the scarcity of natural nesting materials, leading them to resort to readily available anthropogenic substitutes (Wang et al., 2009). Second, the adaptive hypothesis suggests that the selection of ANMs by birds may be related to their increased reproductive performance (Suárez-Rodríguez et al., 2013; Reynolds et al., 2019). Third, the new location hypothesis posits that birds choose different nest materials due to changes in nest sites (Reynolds et al., 2019). Finally, the search image hypothesis indicates that birds using ANMs on the outer layer of the nest will change the original image of the nest and then can avoid predator search (Corrales-Moya et al., 2023).
In this study, we investigated the relationship between the incorporation of ANMs into nests and the reproductive performance of Chinese Bulbuls (Pycnonotus sinensis) by monitoring and collecting data on 136 breeding nests along the urbanization gradient in Hangzhou, China. We aimed to address the following questions: (1) Does urbanization intensity influence the amount of ANMs in the nests of Chinese Bulbuls? (2) Whether ANMs may influence reproductive traits of Chinese Bulbuls?
2.
Materials and methods
2.1
Study area
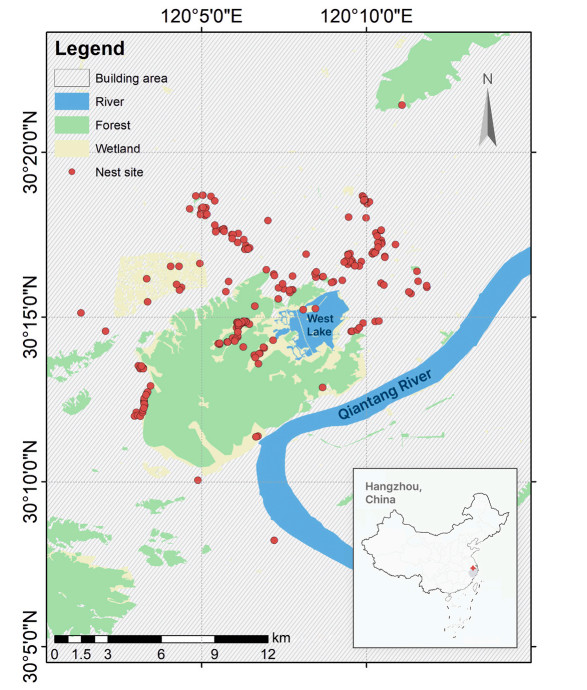
The study was conducted in Hangzhou (29°11′–30°33′ N, 118°21′–120°30′ E) (Fig. 1), the capital city of Zhejiang Province, China. Serving as the administrative, financial and cultural hub of Zhejiang Province, Hangzhou is one the most urbanized cities in the Yangtze River Delta (Wang et al., 2008; Chen et al., 2022). Hangzhou has a population of 12.376 million people as of the year 2022, with a high proportion of urban population of 84.0% (http://www.hangzhou.gov.cn/). The city is covered by hills, flatlands and water systems such as rivers and lakes, which make up 65.6%, 26.4%, and 8.0% of the total land area (Chen et al., 2000), respectively.
Figure
1.
Distribution map of the nest sites of the Chinese Bulbul (Pycnonotus sinensis) in Hangzhou, China.
The Chinese Bulbul is a small passerine bird with widespread distribution across southern China and East Asia (Zhuge, 1990). This bird species is highly adaptable to urban environments and can thrive even in intensely urbanized areas (Wang et al., 2009; Chen et al., 2023). Previous studies have demonstrated that Chinese Bulbuls can adapt to urban ecosystems by altering their nesting materials (Wang et al., 2009), using food provided by humans (Wang, 2003), adjusting their flight initiation distance in urban areas (Wang et al., 2004), and changing reproductive traits such as laying date (Chen et al., 2023). Chinese Bulbuls in urban areas often nest in some tree species such as Osmanthus fragrans, Photinia serrulata, Bambusoideae sp., Elaeocarpus sylvestris (Lan et al., 2013). Chinese Bulbuls build a tall cup nest, which is typically 2–4 m above the ground and usually constructed from natural materials such as hay stems, fine roots, and plant fibers (Zhuge, 1990).
2.3
Nest surveys
During two breeding seasons in 2012 and 2013, we conducted nest surveys across an urbanization gradient in Hangzhou, spanning from intensely constructed urban centers to pristine natural environments of the western mountainous areas (Fig. 1) (Chen et al., 2022, 2023). We searched for active breeding nests of Chinese Bulbuls by listening for their chirping and observing their nesting material transportation behaviors through telescopes (Wang et al., 2009). The detection of bulbuls' nests was further informed by their nest site preferences (Wang, 2003; Chen et al., 2006), favored nesting trees, particularly osmanthus trees (Lan et al., 2013), and observed behaviors of carrying nesting materials (Wang et al., 2009). Once a breeding nest was discovered, we recorded its coordinates using a GPS for subsequent monitoring (Lan et al., 2013). More details about nest surveys have been described in our previous work (Chen et al., 2022).
2.4
Reproductive parameters
We observed the reproductive behaviors of Chinese Bulbuls from mid-March (Lan, 2013). The breeding periods of Chinese Bulbuls were April 9–May 30 in 2012 and April 1–June 4 in 2013 (Lan et al., 2013). We documented seven reproductive parameters, including laying date, incubation period, nestling period, clutch size, egg volume, hatching success rate, and fledging success rate (Chen et al., 2023). The laying date was defined as the day when the first egg was laid. We calculated the incubation duration as the days from the final egg's laying to the first chick's hatching (Chen et al., 2023). The nestling period spanned from the time when the first chick hatched to the fledging date (Ibáñez-Álamo and Soler, 2010). We defined clutch size as the number of eggs in each nest (Bailly et al., 2015). Chinese Bulbuls usually lay one egg per day, and the number of eggs in a clutch is usually 3 or 4 (Zhuge, 1990). Within two days after the end of egg laying, we measured the long and short radii of eggs with Vernier calipers to the millimeter (Lan et al., 2013). Egg volume was then calculated using the formula: Volume (V) = Kv × mean egg length × mean egg width2, in which Kv is the volume coefficient (Hoyt, 1979). All the above measurements were conducted during the periods when parent birds were absent from the nests to reduce potential human disturbance (Zhang, 2014).
We calculated the reproductive success rates for each nest after the end of breeding periods. The hatching success rate was calculated as the number of hatchlings divided by the clutch size (Chen et al., 2023). The fledging success rate was defined as the number of fledglings divided by the number of hatchlings (Corsini et al., 2017).
2.5
Nest composition
Although we initially found 234 natural breeding nests (Chen et al., 2023), after the breeding seasons we eventually obtained 136 nests that can be used for nest material analysis. The other 98 nests were damaged before we removed them from trees due to bad weather conditions and long-time exposures (Zhang, 2014). To maintain the nests' structural integrity and composition, we removed these 136 nests carefully in situ and placed them in well-padded boxes for transportation to our laboratory (Biddle et al., 2019).
Since the collection time was different, we treated each nest in a dryer (HASUC Blower Dryer DHG-9070A) at 60 ℃ for 12 h before performing nest material analysis. We first weighed each nest and then carefully separated all nest materials with clippers (Zhang, 2014). The nesting materials were divided into categories according to the degree of identification (Wang et al., 2009). Finally, we weighed and analyzed each category of nesting materials separately. After dismantling nests of Chinese Bulbuls, we classified all the nest materials into two general groups, i.e. natural or anthropogenic nesting materials (Table 1).
Table
1.
The composition of 136 nests of Chinese Bulbuls in Hangzhou, China. Number of nests represents the total number of nests that contain each type of nesting material, and the proportion represents the ratio of the number of nests divided by the total number of nests (136).
In earlier urbanization research, several simple measures such as the percentage of the urban landscape or the distance to human settlements were used to quantify urbanization intensity (Marzluff et al., 2001). However, these simple measures often overlooked other important urbanization factors like human disturbance and environmental noise (Wang et al., 2008, 2009). Due to the complexities of urban environment, one single factor is not enough to capture the full urbanization context (McDonnell and Pickett, 1990; Wang et al., 2008).
In our study, we measured the following four urbanization characteristic parameters to quantify urbanization intensity (Chen et al., 2022, 2023).
1. Distance to City Center (DCC): Measured as the straight-line distance from the nest site of Chinese Bulbuls to the Hangzhou City Government Hall (the city center of Hangzhou) using the ArcGIS (ArcView 3.3) (Wang et al., 2008).
2. Building Index (BI): Calculated using the ArcGIS (ArcView 3.3) by measuring the proportion of the area covered by buildings around each nest at varying radii (250 m, 500 m, 1000 m, 2000 m) (Bolger et al., 1997; Wang et al., 2008). Building Index was then calculated using the following formula: BI = 250 m × 1 + 500 m × 0.5 + 1000 m × 0.25 + 2000 m × 0.125.
3. Environmental Noise (EN): Measured using a HS5633A sound meter near the nest in 3-min intervals from 09:00 to 10:00 h on both weekdays and weekends. The average environmental noise (dB) was calculated as: EN = weekday noise × 5/7 + weekend noise × 2/7 (Chen et al., 2022, 2023).
4. Disturbance Index (DI): Represents the number of people and vehicles passing by the nest between 8:00 to 10:00 a.m. It was the sum of average foot traffic and vehicle traffic, both of which were calculated similarly to the environmental noise (Chen et al., 2022, 2023).
These four urbanization characteristic parameters were standardized to values between 0 and 100 and then were combined to create the Urbanization Synthetic Index (USI) to quantitively measure urbanization intensity as follows (Wang et al., 2008; Chen et al., 2022): USI = BI × 100/2 + EN + DI/3 + 1/DCC × 100.
2.7
Data analyses
For each nest, we measured the weight of ANMs and the proportion of ANMs (the weight of ANMs/total weight of the nest). We used the arcsine square root transformation to convert the proportion of ANMs (Zar, 1999) and log-transformed the USI. Prior to analysis, we used the R package "fitdistrplus" (Delignette-Muller and Dutang, 2015) to examine the distribution of the dependent variable. We then constructed a linear mixed effect model (LMM) (Bates et al., 2015) to examine the relationship between the USI and the amount of ANMs, with USI as the fixed effect and tree species as a random effect (Chen et al., 2023).
We applied binary logistic regressions to analyze the relationship between the amount of ANMs and clutch size. We only observed three or four eggs in the nests of Chinese Bulbuls after removing invalid data such as abandoned nests or predated nests (Chen et al., 2023). Therefore, we converted clutch size into a two-valued response variable (three eggs = 0, four eggs = 1) and then applied binary logistic regressions to analyze its relationship with the proportion and weight of ANMs.
We built LMMs to analyze the influence of the amount of ANMs on the other six reproductive parameters (laying date, incubation period, nestling period, egg volume, hatching success rate, and fledging success rate). In these LMMs, we used the proportion or weight of ANMs as the fixed effect and tree species as random factor. Before data analyses, we converted the hatching success rate and fledging success rate using the arcsine square root transformation.
Values are given as mean ± SE, all tests are two-tailed, and statistical significance was set at α ≤ 0.05 (Zar, 1999). All analyses were performed in R 4.1.2 (R Core Team, 2021).
3.
Results
Among the 136 examined nests of Chinese Bulbuls, 131 (96.32%) contained anthropogenic materials. Among the types of ANMs, the most highest frequency was plastic (95.6%) and chemical fiber (89.0%), while the lowest frequency was metal (0.7%). Among natural nesting materials, stems and leaves were the most frequent (98.5%), whereas feathers (0.7%) were the lowest (Table 1).
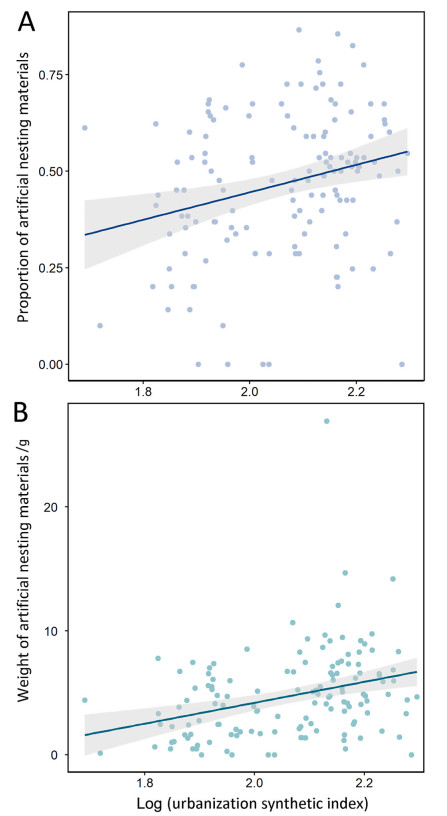
Both the proportion and weight of ANMs in the nests of Chinese Bulbuls increased significantly with the urbanization synthetic index (Fig. 2; t1 = 3.463, p < 0.01; t2 = 4.220, p < 0.01).
Figure
2.
The relationship between the content of anthropogenic nesting materials in nests of Chinese Bulbuls and the degree of urbanization. (A: proportion of anthropogenic nesting materials, arcsine square root; B: weight of anthropogenic nesting materials).
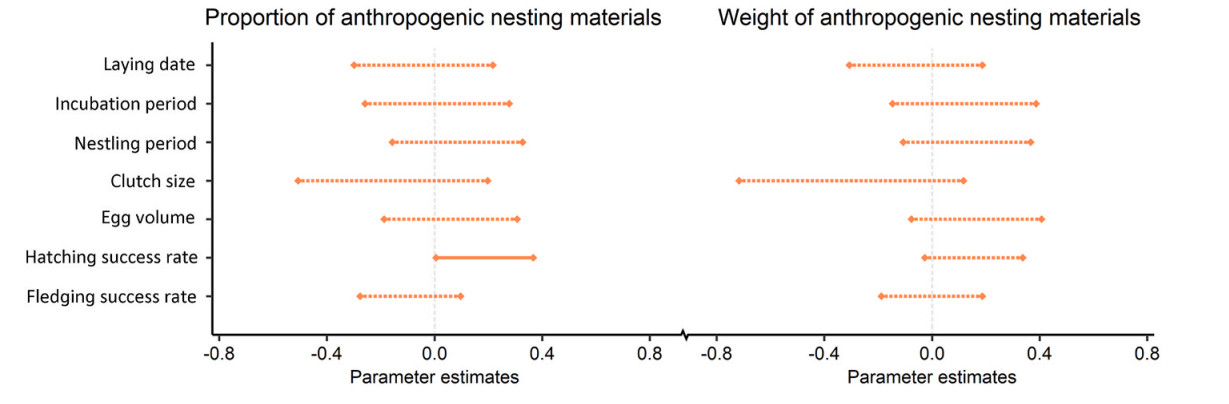
We observed a significantly higher hatching success rate for Chinese Bulbuls with an increasing proportion of ANMs (Fig. 3, Table 2). However, the other six reproductive traits, i.e., laying date, incubation period, nestling period, clutch size, egg volume, and fledging success rate, did not vary significantly with the proportion of ANMs (Table 2). In addition, the weight of ANMs was not significantly correlated with all the seven reproductive traits (Table 3).
Figure
3.
The relationship between the content of anthropogenic nesting materials and reproductive traits of Chinese Bulbuls. Solid lines represent significant influences, based on whether the 95% CI overlaps zeroes.
Table
2.
Results of linear mixed effects models (LMMs) and binary logistic regressions relating the proportion of anthropogenic nesting materials to reproductive traits of Chinese Bulbuls. n is the sample size for each variable. Significant results are shown in bold type.
Table
3.
Results of linear mixed effects models (LMMs) and binary logistic regressions relating the weight of anthropogenic nesting materials to reproductive traits of Chinese Bulbuls. n is the sample size for each variable.
In this study, we systematically examined the relationship between the incorporation of ANMs into nests and the reproductive performance of Chinese Bulbuls by monitoring and collecting data on 136 breeding nests in Hangzhou, China. We found significant increases in both the proportion and weight of ANMs in the nests of Chinese Bulbuls with increasing urbanization intensity. This result may be related to the reduction of natural nesting material in urbanized environment (Wang et al., 2009). With the advancement of urbanization, the natural environment and ecosystems have undergone significant changes. Natural habitats such as forests and grasslands are gradually replaced by artificial facilities such as buildings and roads (Marzluff et al., 2001). Many natural resources are exploited and utilized by birds, resulting in a scarcity of sources of natural nest timber (e.g., twigs, dead grass stems and leaves, etc.). These results indicate that as one of the urban adaptable species, Chinese Bulbuls choose to use ANMs for nesting in highly urbanized habitats.
Our analysis shows that the Chinese Bulbul's hatching success rate is significantly improved after adding ANMs to the nests. The success of bird hatching is closely related to the right temperature (Deeming and Pike, 2015). A stable temperature is essential for embryonic development (Lundy, 1969). As some kinds of ANMs, such as cotton, can better keep the temperature in the nest stable, which in turn may improve the hatching success rate of Chinese Bulbuls. In our study, we found that the proportion of all nests containing cotton reached 40.4% (Table 1). It should be noted that a thermoregulation experiment is needed to prove the exact role of ANMs (e.g., cotton) in providing insulation properties in nests.
Our result appears to be consistent with the adaptive hypothesis (Reynolds et al., 2019), which argues that the inclusion of ANMs in the nest is consistent with the reproductive advantage of bird species. However, our findings do not fully support the adaptive hypothesis, as adding ANMs to the nest did not influence other reproductive traits of the Chinese Bulbul. Several studies also have found the potential hazards of ANMs for other bird species. For instance, plastics in nests might ensnare or harm chicks, even proving fatal (Montevecchi, 1991; Baker et al., 2002; Votier et al., 2011). There is also the danger of birds inadvertently feeding on debris, which can decrease their reproductive success (Mee et al., 2007; Kim et al., 2023). Therefore, the benefits and pitfalls of ANMs are quite complex and need more comprehensive exploration. Our result also accords with the availability hypothesis, because our previous study (Wang et al., 2009) found that in land types with different levels of urbanization, the degree of ANMs used in the nests of Chinese Bulbuls increased significantly with the availability of ANMs in the environments. However, the new location hypothesis is not supported, since Chinese bulbuls in our study always nest in trees or shrubs and do not change their nesting locations (Wang et al., 2009; Lan et al., 2013). Finally, we cannot test the search image hypothesis due to the lack of data on the proportion of nest area covered with ANMs.
The results showed that the proportion or weight of ANMs was not correlated with the other reproductive traits of Chinese Bulbuls. Several reasons may explain why some changes of reproductive traits with ANMs are not noticeable. First, it may be difficult for ANMs to influence certain relatively fixed reproductive traits of Chinese Bulbuls, such as the clutch size (either three or four) or egg volume (Zhang, 2014). Second, the sample sizes for all the reproductive traits among the 136 breeding nests was not consistent due to several uncontrollable or logistical reasons, such as bad weathers, abandoned nests, and labor shortage (Chen et al., 2023). The inconsistence in sample sizes may also lead to the weak relationship between ANMs and reproductive traits of Chinese Bulbuls. Finally, in previous studies, the use of ANMs has adversely affected the physical condition of chicks and adults (Suárez-Rodríguez et al., 2014; 2017), indicating that the effects of ANMs on birds may be manifested in subsequent stages following the breeding period.
5.
Conclusion
In conclusion, we found that Chinese Bulbuls choose ANMs to build their nests, with 96.32% of examined nests containing such anthropogenic components. Our analysis showed that the incorporation of ANMs into nests significantly increased the hatching success rate of Chinese Bulbuls, but was not correlated with other reproductive traits. Overall, the use of ANMs by Chinese Bulbuls seems to be consistent with the adaptive hypothesis and the availability hypothesis. However, it should be noted that our results and conclusions on the relationship between the incorporation of ANMs into nests and the reproductive performance of Chinese Bulbuls are primarily based on field observations. Further studies should use controlled experiments to accurately investigate the impact of ANMs on reproductive success.
Ethics statement
Our study was carried out in agreement with the current law of China on the protection of wildlife.
CRediT authorship contribution statement
Xingmin Chen: Writing – original draft, Validation, Software, Methodology, Formal analysis, Data curation. Qin Zhang: Validation, Methodology, Investigation, Data curation. Sisi Lan: Validation, Methodology, Investigation, Data curation. Qin Huang: Validation, Methodology, Investigation, Data curation. Shuihua Chen: Supervision, Resources, Project administration, Methodology, Investigation, Funding acquisition, Data curation, Conceptualization. Yanping Wang: Writing – review & editing, Supervision, Resources, Project administration, Methodology, Funding acquisition, Conceptualization.
Declaration of competing interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Acknowledgements
We thank Tengda Zhu, Yafei Ma, Jianwei Zhou, Chao Chen, Xiang Yu, Wen Xu, Mingming Hu and Chenchen Zhu for their field assistance in finding and monitoring bird nests.
Bailly, J., Scheifler, R., Berthe, S., Clément-Demange, V., Leblond, M., Pasteur, B., et al., 2015. From eggs to fledging: negative impact of urban habitat on reproduction in two tit species. J. Ornithol. 157, 377-392 .
Baker, G.B., Rosemary, G., Hamilton, S., Wilkinson, V., 2002. Albatrosses and petrels in Australia: a review of their conservation and management. Emu 102, 71-97 .
Biddle, L.E., Dickinson, A.M., Broughton, R.E., Gray, L.A., Bennett, S.L., Goodman, A.M., et al., 2019. Construction materials affect the hydrological properties of bird nests. J. Zool. 309, 161-171 .
Bolger, D.T., Scott, T.A., Rotenberry, J.T., 1997. Breeding bird abundance in an urbanizing landscape in coastal southern California. Conserv. Biol. 11, 406-421 .
Bates, D.M., Mächler, M., Bolker, B., Walker, S., 2015. Fitting linear mixed-effects models using lme4. J. Stat. Software 67, 1-48.
Briggs, K.B., Deeming, D.C., Mainwaring, M.C., 2023. Plastic is a widely used and selectively chosen nesting material for pied flycatchers (Ficedula hypoleuca) in rural woodland habitats. Sci. Total Environ. 854, 158660 .
Chen, S., Ding, P., Zheng, G., Zhuge, Y., 2000. Effects of urbanization on wetland waterbird communities in Hangzhou. Zool. Res. 21, 279-285.
Chen, W., Wu, Y., Hu, J., Yu, Z., Guo, Z., 2006. Nest site selection and materials of Pycnonotus sinensis in Nanchong, Sichuan. Sichuan J. Zool. 25, 590-593.
Chen, X., Zhang, Q., Lan, S., Chen, S., Wang, Y., 2022. Nest predation pressure on Chinese Bulbuls decreases along the urbanization gradient in Hangzhou, China. Avian Res. 13, 100049 .
Chen, X., Zhang, Q., Lan, S., Chen, S., Wang, Y., 2023. Variation in reproductive life-history traits of Chinese Bulbuls (Pycnonotus sinensis) along the urbanization gradient in Hangzhou, China. Avian Res. 14, 100100 .
Collias, N.E., Collias, E.C., 2016. Nest building and bird behavior. Princeton University Press, New Jersey.
Corrales-Moya, J., Barrantes, G., Chacón-Madrigal, E., Sandoval, L., 2021. Human waste used as nesting material affects nest cooling in the clay-colored thrush. Environ. Pollut. 284, 117539 .
Corrales-Moya, J., Barrantes, G., Chacón-Madrigal, E., Sandoval, L., 2023. A potential consequence for urban birds' fitness: exposed anthropogenic nest materials reduce nest survival in the clay-colored thrush. Environ. Pollut. 326, 121456 .
Corsini, M., Dubiec, A., Marrot, P., Szulkin, M., 2017. Humans and tits in the city: quantifying the effects of human presence on great tit and blue tit reproductive trait variation. Front. Ecol. Evol. 5, 82 .
Deeming, D.C., Pike, T.W., 2015. Nest surface temperature predicts fledging success of Blue Tits Cyanistes caeruleus but not Great Tits Parus major. Acta Ornithol 50, 247-251 .
Deeming, D.C., Reynolds, S.J., 2016. Nests, eggs, and incubation: New ideas about avian reproduction. Oxford University Press, Oxford.
Delignette-Muller, M.L., Dutang, C., 2015. Fitdistrplus: an R package for fitting distributions. J. Stat. Software 64, 1-34 .
Devictor, V., Julliard, R., Couvet, D., Lee, A., Jiguet, F., 2007. Functional homogenization effect of urbanization on bird communities. Conserv. Biol. 21, 741-751 . PMID: 17531052.
Evans, K.L., Gaston, K.J., Frantz, A.C., Simeoni, M., Sharp, S.P., McGowan, A., et al., 2009. Independent colonization of multiple urban centres by a formerly forest specialist bird species. Proc. R. Soc. B 276, 2403-2410 .
Hansell, M.H., 2005. Bird nests and construction behaviour. Cambridge University Press, Cambridge.
Hoyt, D.F., 1979. Practical methods of estimating volume and fresh weight of bird eggs. Auk 96, 73-77 .
Ibáñez-Álamo, J.D., Soler, M., 2010. Does urbanization affect selective pressures and life-history strategies in the common blackbird (Turdus merula L.)? Biol. J. Linn. Soc. 101, 759-766 .
Jagiello, Z., Corsini, M., Dylewski, Ł., Ibáñez-Álamo, J.D., Szulkin, M., 2022. The extended avian urban phenotype: anthropogenic solid waste pollution, nest design, and fitness. Sci. Total Environ. 838, 156034 .
Jagiello, Z., Dylewski, Ł., Tobolka, M., Aguirre, J.I., 2019. Life in a polluted world: a global review of anthropogenic materials in bird nests. Environ. Pollut. 251, 717-722 .
Jagiello, Z., Reynolds, S.J., Nagy, J., Mainwaring, M.C., Ibáñez-Álamo, J.D., 2023. Why do some bird species incorporate more anthropogenic materials into their nests than others? Phil. Trans. R. Soc. B 378, 20220156 .
Kaza, S., Yao, L.C., Bhada-Tata, P., van Woerden, F., 2018. What a Waste 2.0: A Global Snapshot of Solid Waste Management to 2050. Urban Development. World Bank, Washington, DC.
Kim, M., Hong, M., Nam, K., Kim, Y., Park, C., Kwon, Y., 2023. Marine debris ingestion by adults and fledglings of Swinhoe's storm petrels in the Republic of Korea. Mar. Pollut. Bull. 194, 115330 .
Lan, S., 2013. The breeding ecology of Chinese Bulbul and it's responses of breeding behavior to urbanization in Hangzhou, China. Master's Thesis. Zhejiang Normal University, Jinhua.
Lan, S., Zhang, Q., Huang, Q., Chen, S., 2013. Breeding ecology of Chinese Bulbul in the urban environment of Hangzhou, China. Zool. Res. 34, 182-189.
Lavers, J.L., Bond, A.L., Hutton, I., 2014. Plastic ingestion by Flesh-footed Shearwaters (Puffinus carneipes): implications for fledgling body condition and the accumulation of plastic-derived chemicals. Environ. Pollut. 187, 124-129 .
Leveau, L.M., Isla, F.I., Bellocq, M.I., 2015. Urbanization and the temporal homogenization of bird communities: a case study in central Argentina. Urban Ecosyst. 18, 1461-1476 .
Lundy, H., 1969. A review of the effects of temperature, humidity, turning and gascous
environment in the incubator on the hatchability of the hen's egg. In: Carter, T.C.,
Freeman, B.M. (Eds.), The Fertility and Hatchability of the Hen's Egg. Oliver and
Boyd, Edinburgh, Scotland, pp. 143–176.
Mainwaring, M.C., 2015. The use of man-made structures as nesting sites by birds: a review of the costs and benefits. J. Nat. Conserv. 25, 17-22 .
Mansfield, I., Reynolds, S.J., Lynch, I., Matthews, T.J., Sadler, J.P., 2024. Birds as bioindicators of plastic pollution in terrestrial and freshwater environments: a 30-year review. Environ. Pollut. 348, 123790 .
Marzluff, J.M., Bowman, R., Donelly, R., 2001. Avian Ecology and Conservation in an Urbanizing World. Kluwer Academic Publishers, Boston.
McDonnell, M.J., Pickett, S.T.A., 1990. Ecosystem structure and function along urban–rural gradients: an unexploited opportunity for ecology. Ecology 71, 1232-1237 .
Mee, A., Rideout, B.A., Hamber, J.A., Todd, N., Austin, G., Clark, M., et al., 2007. Junk ingestion and nestling mortality in a reintroduced population of California Condors Gymnogyps californianus. Bird. Conserv. Int. 17, 119-130 .
Minias, P., 2015. Successful colonization of a novel urban environment is associated with an urban behavioural syndrome in a reed-nesting waterbird. Ethology 121, 1178-1190 .
Morelli, F., Mikula, P., Benedetti, Y., Bussière, R., Jerzak, L., Tryjanowski, P., 2018. Escape behaviour of birds in urban parks and cemeteries across Europe: evidence of behavioural adaptation to human activity. Sci. Total Environ. 631–632, 803-810 .
Montevecchi, W.A., 1991. Incidence and types of plastic in gannets' nests in the northwest Atlantic. Can. J. Zool. 69, 295-297 .
Montoya-Tangarife, C., de la Barrera, F., Salazar, A., Inostroza, L., 2017. Monitoring the effects of land cover change on the supply of ecosystem services in an urban region: a study of Santiago-Valparaíso, Chile. PLoS One 12, e0188117 .
R Core Team, 2021. R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna. .
Reynolds, S.J., Ibáñez-Álamo, J.D., Sumasgutner, P., Mainwaring, M.C., 2019. Urbanisation and nest building in birds: a review of threats and opportunities. J. Ornithol. 160, 841-860 .
Setälä, O., Fleming-lehtinen, V., Lehtiniemi, M., 2014. Ingestion and transfer of microplastics in the planktonic food web. Environ. Pollut. 185, 77-83 .
Seto, K.C., Güneralp, B., Hutyra, L.R., 2012. Global forecasts of urban expansion to 2030 and direct impacts on biodiversity and carbon pools. P. Natl. Acad. Sci. USA 109, 16083-16088 .
Sheard, C., Stott, L., Street, S.E., Healy, S.D., Sugasawa, S., Lala, K.N., 2024. Anthropogenic nest material use in a global sample of birds. J. Anim. Ecol. 93, 691-704 .
Suárez-Rodríguez, M., Garcia, M., 2014. There is no such a thing as a free cigarette; lining nests with discarded butts brings short-term benefits, but causes toxic damage. J. Evol. Biol. 27, 2719-2726 .
Suárez-Rodríguez, M., López-Rull, I., Macías Garcia, C., 2013. Incorporation of cigarette butts into nests reduces nest ectoparasite load in urban birds: new ingredients for an old recipe? Biol. Lett. 9, 20120931 .
Suárez-Rodríguez, M., Montero-Montoya, R.D., Macías Garcia, C., 2017. Anthropogenic nest materials may increase breeding costs for urban birds. Front. Ecol. Evol. 5. .
UNEP, 2014. UNEP Year Book 2014 Emerging Issues Update. United Nations Environment Programme, Nairobi, Kenya.
Votier, S.C., Archibald, K., Morgan, G., Morgan, L., 2011. The use of plastic debris as nesting material by a colonial seabird and associated entanglement mortality. Mar. Pollut. Bull. 62, 168-172 .
Wang, Y., 2003. A Study of the Adaptability of Birds to Urbanization. Master's Thesis. Zhejiang University, Hangzhou.
Wang, Y., Chen, S., Blair, R.B., Jiang, P., Ding, P., 2009. Nest composition adjustments by Chinese bulbuls (Pycnonotus sinensis) in an urbanized landscape of Hangzhou (E China). Acta Ornithol. 44, 185–192. (Briggs).
Wang, Y., Chen, S., Ding, P., 2004. Flush distance: birds tolerance to human intrusion in Hangzhou. Zool. Res. 25, 214-220.
Wang, Y., Chen, S., Jiang, P., Ding, P., 2008. Black-billed Magpies (Pica Pica) adjust nest characteristics to adapt to urbanization in Hangzhou, China. Can. J. Zool. 86, 676-684 .
Wang, Y., Huang, Q., Zhang, Q., Lan, S., Chen, S., 2015. Common blackbirds Turdus merula use anthropogenic structures as nesting sites in an urbanized landscape. Curr. Zool. 61, 435-443.
Zar, J.H., 1999. Biostatistic Analysis, fourth ed. Prentice Hall Inc., Upper Saddle River, New Jersey.
Zhang, Q., 2014. Responses of Life History Traits of Chinese Bulbul to Urbanization. Master's Thesis. Zhejiang Normal University, Jinhua.
Zhuge, Y., 1990. Fauna of Zhejiang: Aves. Zhejiang Science and Technology Press, Hangzhou China.
Shuyu Hou, Rui Yang, Yue Cao, et al. A framework for identifying bird conservation priority areas in croplands at national level. Journal of Environmental Management, 2022, 324: 116330.
DOI:10.1016/j.jenvman.2022.116330
Table
1.
The composition of 136 nests of Chinese Bulbuls in Hangzhou, China. Number of nests represents the total number of nests that contain each type of nesting material, and the proportion represents the ratio of the number of nests divided by the total number of nests (136).
Table
2.
Results of linear mixed effects models (LMMs) and binary logistic regressions relating the proportion of anthropogenic nesting materials to reproductive traits of Chinese Bulbuls. n is the sample size for each variable. Significant results are shown in bold type.
Table
3.
Results of linear mixed effects models (LMMs) and binary logistic regressions relating the weight of anthropogenic nesting materials to reproductive traits of Chinese Bulbuls. n is the sample size for each variable.

 DownLoad:
DownLoad:





 Email Alerts
Email Alerts RSS Feeds
RSS Feeds