Kui Yan, Wei Liang. 2024: Recognition and rejection of foreign eggs of different colors in Barn Swallows. Avian Research, 15(1): 100202. DOI: 10.1016/j.avrs.2024.100202
Citation:
Kui Yan, Wei Liang. 2024: Recognition and rejection of foreign eggs of different colors in Barn Swallows. Avian Research, 15(1): 100202. DOI: 10.1016/j.avrs.2024.100202
Kui Yan, Wei Liang. 2024: Recognition and rejection of foreign eggs of different colors in Barn Swallows. Avian Research, 15(1): 100202. DOI: 10.1016/j.avrs.2024.100202
Citation:
Kui Yan, Wei Liang. 2024: Recognition and rejection of foreign eggs of different colors in Barn Swallows. Avian Research, 15(1): 100202. DOI: 10.1016/j.avrs.2024.100202
Ministry of Education Key Laboratory for Ecology of Tropical Islands, Key Laboratory of Tropical Animal and Plant Ecology of Hainan Province, College of Life Sciences, Hainan Normal University, Haikou, 571158, China
Funds:
the National Natural Science Foundation of China31970427
the National Natural Science Foundation of China32270526 to W.L.
Brood parasitic birds lay eggs in the nests of other birds, and the parasitized hosts can reduce the cost of raising unrelated offspring through the recognition of parasitic eggs. Hosts can adopt vision-based cognitive mechanisms to recognize foreign eggs by comparing the colors of foreign and host eggs. However, there is currently no uniform conclusion as to whether this comparison involves the single or multiple threshold decision rules. In this study, we tested both hypotheses by adding model eggs of different colors to the nests of Barn Swallows (Hirundo rustica) of two geographical populations breeding in Hainan and Heilongjiang Provinces in China. Results showed that Barn Swallows rejected more white model eggs (moderate mimetic to their own eggs) and blue model eggs (highly non-mimetic eggs with shorter reflectance spectrum) than red model eggs (highly non-mimetic eggs with longer reflectance spectrum). There was no difference in the rejection rate of model eggs between the two populations of Barn Swallows, and clutch size was not a factor affecting egg recognition. Our results are consistent with the single rejection threshold model. This study provides strong experimental evidence that the color of model eggs can has an important effect on egg recognition in Barn Swallows, opening up new avenues to uncover the evolution of cuckoo egg mimicry and explore the cognitive mechanisms underlying the visual recognition of foreign eggs by hosts.
Avian brood parasitism is the process by which parasitic birds lay their eggs in the nests of other species (hosts), tricking the host into raising the unrelated offspring. Visual recognition and rejection of foreign eggs is a common antiparasitic defense method adopted by many hosts (Rothstein, 1990; Lahti, 2006; Soler, 2014; Manna et al., 2017). A popular explanation for the cognitive mechanisms underlying host recognition of foreign eggs has long assumed that the degree of difference between the background colors of foreign and host eggshells (that is absolute color contrast) is the strongest linear predictor of egg recognition (Avilés, 2008; Cassey et al., 2008; Stoddard and Stevens, 2011; Stevens et al., 2013; Hauber et al., 2015; Manna et al., 2019). This implies that a host with an egg color of mid-wavelength visible light can identify and reject foreign eggs with egg color wavelengths that are significantly shorter (bluer) and longer (redder) than their own through color comparison (colors in parentheses are based on human perception). However, recent studies have shown that birds have nonlinear egg color perception (Caves et al., 2018; Zipple et al., 2019), and nonlinear sensory and cognitive processes may also contribute to or fully mediate the decision of parasitized avian hosts to recognize foreign eggs.
Recent studies investigating the responses of two host species to model parasitic eggs along the natural egg color gradient of birds have revealed a new cognitive decision rule (Hanley et al., 2015, 2017), which has since been repeatedly confirmed (Abolins-Abols et al., 2019; Hanley et al., 2019). These cognitive patterns have been documented in European Blackbirds (Turdus merula) in the Czech Republic, American Robins (T. migratorius) of the same genus in the USA (Hanley et al., 2017), and Chalk-browed Mockingbirds (Mimus saturninus) in Argentina (Hanley et al., 2019). These host species breed on different continents and are parasitized by different parasitic species. Similar experimental results have been observed in Great Reed Warblers (Acrocephalus arundinaceus) in Hungary (Abolins-Abols et al., 2019), which was the first among these host species tested where the parasitic eggs closely mimicked host eggs (Igic et al., 2012). When analyzed using ultraviolet-sensitive avian visual modeling parameters, all birds in these studies preferentially rejected foreign eggs that were redder than their own egg color and preferentially accepted foreign eggs that were bluer than their own egg color. These findings imply that there is a single color-based threshold for egg recognition that exists at some distance from the host's own egg color to a redder color, rather than the multiple rejection thresholds as assumed by previous theories (Reeve, 1989; Hauber et al., 2006).
The Barn Swallow (Hirundo rustica) is a common insectivorous bird widely distributed worldwide, mostly breeding in temperate regions and migrating to the tropics or the Southern Hemisphere to overwinter (Turner, 2006). Compared to temperate, even Barn Swallows breeding in the most southern Chinese mainland also exhibited migratory behaviour, and wintering on various islands in Southeast Asia (Tian et al., 2024). The Barn Swallow has open cup-shaped nests made out of mud, straw and feathers, having a clutch of 3–5 eggs, with spotted white eggs of 18.41 ± 0.91 mm × 13.27 ± 0.44 mm in egg size and 1.51 ± 0.23 g in egg mass (Yang et al., 2015b). The extensive breeding range of the Barn Swallow is believed to be due in part to their close association with human populations, and this association with humans has persisted for millennia (Zink et al., 2006). Barn Swallows are thus well known and easily recognizable throughout rural areas worldwide (Scordato and Safran, 2014) but serves as a rare host for the Common Cuckoo (Cuculus canorus), with a parasitism rate of 2.4% (n = 42) in Jilin, northeastern China, and additional 13 cases of parasitism, all in northern China (Su et al., 2017). Campobello and Sealy (2009) reported a parasitism rate of 1.2% (n = 416 nests) in Italy, and parasitism rate was 0.2% (n = 1530) in the Polish nest record scheme (Liang et al., 2013). Barn Swallows showed a moderate egg rejection rate towards non-mimetic eggs but almost accept mimetic eggs. For example, Barn Swallows rejected 24% of blue model eggs in northern China (n = 17), 50% in southern China (n = 18) (Yang et al., 2015b), and 49.2% in Hainan Island (n = 63; Yang et al., 2022a), while the rejection rate was 63% in Denmark (n = 27; Yang et al., 2015b). In Czech Republic, Barn Swallows rejected 29.6% of blue model eggs (n = 27) but only 3.6% of mimetic white spotted model eggs (n = 28; Šulc et al., 2022). It seemed that egg rejection of Barn Swallows varied with geographic populations and different colors of eggs. In this study, we examined the parasitized Barn Swallow population in Heilongjiang Province in northeastern China and the non-parasitized population in Hainan Province in southern China (Su et al., 2017). If the single rejection threshold model applies to Barn Swallows, then we expect the host response to depend on the direction of the color change, not merely the absolute color difference between the foreign eggs and their own eggs. Otherwise, Barn Swallows will reject foreign eggs based on absolute color contrast differences, independent of the direction of color difference. This type of selection may also exist because previous tests of egg recognition in Barn Swallows found that they displayed good recognition for blue model eggs (Yang et al., 2015a, 2015b, 2022a), which contrasts with the species described previously that are more biased toward recognizing brown and accepting blue foreign eggs (Hanley et al., 2015, 2017, 2019; Abolins-Abols et al., 2019). Therefore, Barn Swallows may accept foreign eggs with a mimetic egg color (e.g. white) and reject those that are bluer or redder than the egg color of their own species. Finally, if the frequency of interactions with brood parasites determines the evolution of recognition thresholds for a given host (Reeve, 1989), then we predict that egg recognition thresholds of Barn Swallows are narrower and more stringent in Heilongjiang (parasitized) than in Hainan (non-parasitized) populations.
2.
Materials and methods
2.1
Study area
The three study sites were located in Ding'an (19°13′–19°44′ N, 110°7′–110°31′ E) and Danzhou Counties (19°11′–19°52′ N, 108°56′–109°46′ E) of Hainan, southern China, and Zhalong National Nature Reserve in Qiqihar (46°48′–47°31′ N, 123°51′–124°37′ E) of Heilongjiang, northeastern China. Hainan Island is located at the edge of the low-latitude tropics and has a tropical oceanic monsoon climate, whereas the Zhalong National Nature Reserve has a temperate continental monsoon alpine-lake climate (Yang et al., 2015a, 2019, 2022a).
2.2
Model egg experiments
The experiments had been carried out during the year 2020 for Heilongjiang and Danzhou, Hainan, the year 2021 for Ding'an, Hainan. Once the nests were found, the locations and clutch sizes of the nests were recorded. In addition, nest illuminance of experimental nests was measured by using an ST-80C illuminometer (lux as unit; Photoelectric Instrument Factory of Beijing Normal University, China) (see also Yang et al., 2022b). Model eggs were constructed using clay with reference to the method described by Canniff et al. (2018), and the behavioral responses of Barn Swallows to three different egg colors, that is, white model eggs similar to their own eggs (moderate egg mimicry, Fig. 1a), blue model eggs with shorter wavelengths, and red model eggs with longer wavelengths (both blue and red model eggs were highly non-mimetic, Fig. 1b and c), were tested. All experimental model eggs used were similar to eggs of the Barn Swallow host in egg mass and size. A white, blue, or red model egg was placed into a Barn Swallow nest during incubation period (Fig. 1), and the nest was checked within six days to observe whether the model egg had been recognized and ejected by the Barn Swallows. The responses of Barn Swallows were categorized as either rejection (model eggs ejected from the nest) or acceptance (model eggs found in the nest at the time of examination), and abandoned and predated nests were excluded from the analysis (Yang et al., 2022a). The egg recognition experiment was performed only once for each nest.
Figure
1.
Nests and eggs of Barn Swallows and the added experimental eggs of three different colors (A: the white model egg, B: the blue model egg, C: the red model egg). (For interpretation of the references to color in this figure legend, the reader is referred to the Web version of this article.)
We used a Generalized Linear Mixed Model (GLMM), generated using the glmmTMB package in R version 4.1.3, to analyze the response (accepted or rejected) toward model egg across different colors (blue vs. red vs. white) and clutch size when the model egg was first added to the nest in different study sites (Zhalong vs. Danzhou vs. Ding'an) by Barn Swallows. In this model, the response (accepted or rejected) was the dependent variable. The model egg color, site, nest illuminance, clutch size and the interacting effect of model egg color and different sites were considered as fixed effects. The nest ID was treated as a random effect. Then we used multcomp and emmeans packages for multiple comparative analysis. All tests were two-tailed, and significance levels were all set at P < 0.05.
3.
Results
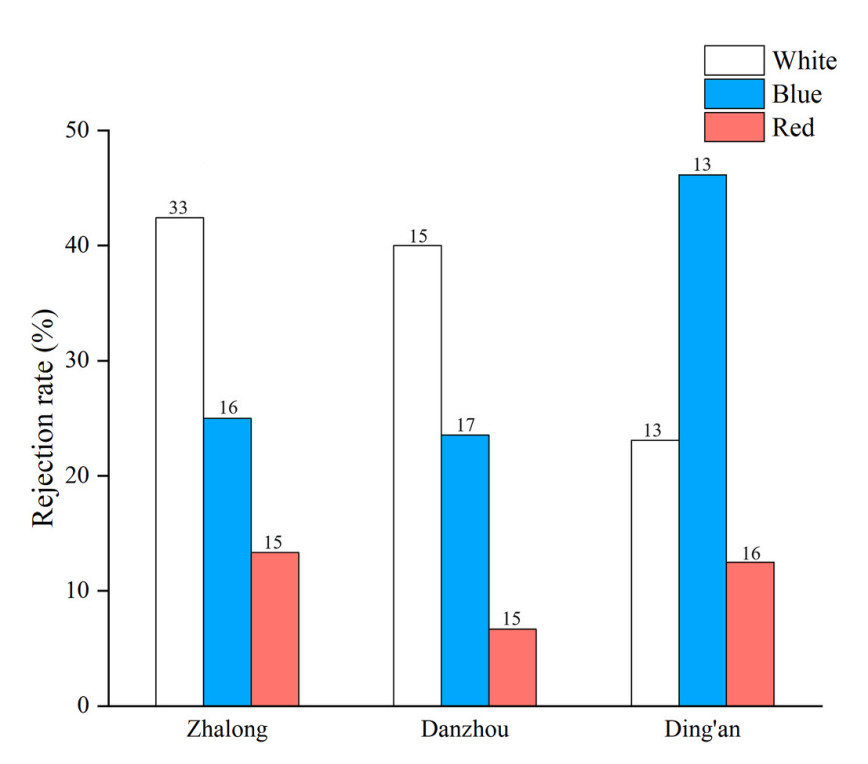
In this study, a total of 153 valid experiments of egg recognition were recorded, including 47 in Danzhou (15 nests for white model eggs, 17 nests for blue model eggs and 15 nests for red model eggs, respectively), 42 in Ding'an (13 nests for white model eggs, 13 nests for blue model eggs and 16 nests for red model eggs, respectively) and 64 in Zhalong (33 nests for white model eggs, 16 nests for blue model eggs and 15 nests for red model eggs, respectively).
The analysis of GLMMs revealed that only model egg color (χ2 = 6.569, P = 0.037) significantly predicted the host response to model eggs (Table 1). The egg rejection of Barn Swallows at three study sites (23.4% in Danzhou, n = 47; 26.2% in Ding'an, n = 42; 31.3% in Zhalong, n = 64) did not show a significant difference (χ2 = 1.335, P = 0.513) (Fig. 2). Additionally, none of the clutch sizes (χ2 = 1.803, P = 0.179), the light intensity (χ2 = 1.857, P = 0.173) or the interaction between experimental egg color and study site (χ2 = 2.801, P = 0.592) had a significant impact on the egg rejection of Barn Swallows (Table 1).
Table
1.
Outcomes of generalized linear mixed-effects model (GLMM) to investigate response to model eggs of different colors by Barn Swallows.
Figure
2.
Egg rejection of Barn Swallows with three different egg colors at three study sites. Number on top of each bar refers to sample size (nests tested). (For interpretation of the references to color in this figure legend, the reader is referred to the Web version of this article.)
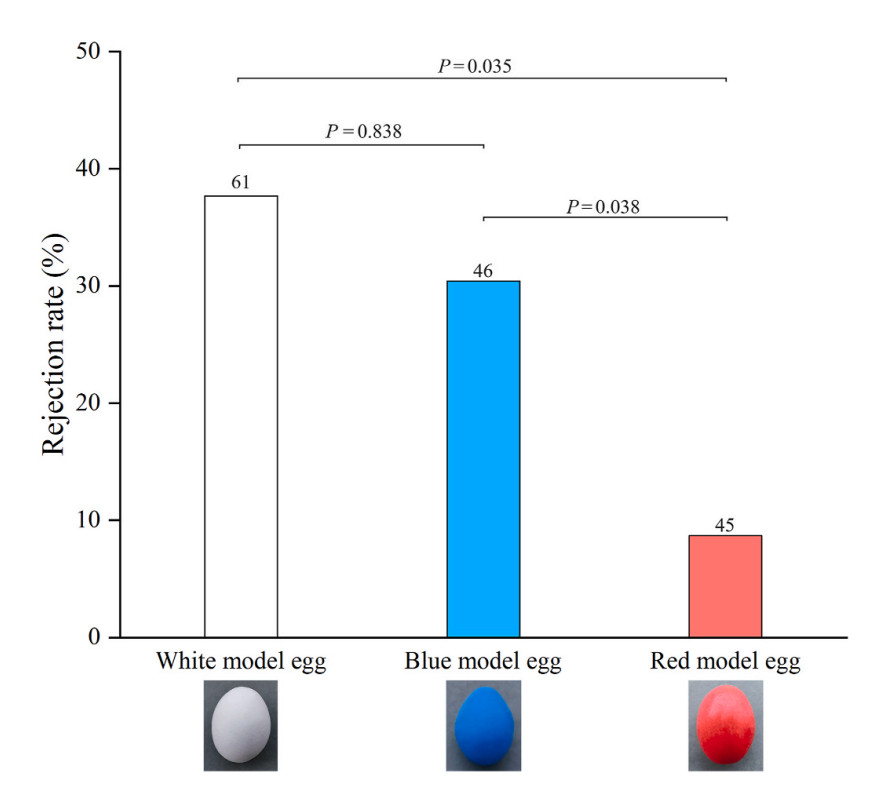
Further analysis for post hoc comparison indicated that the rejection rate of red model eggs by Barn Swallows was significantly lower than that of white model eggs (Z = −2.518, P = 0.035) and blue model eggs (Z = −2.350, P = 0.038). However, there was no significant difference in the rejection rates of white and blue model eggs by Barn Swallows (Z = 0.204, P = 0.838) (Fig. 3).
Figure
3.
Egg rejection rates of Barn Swallows with three different egg colors. Number on top of the bar refers to sample size (nests tested). (For interpretation of the references to color in this figure legend, the reader is referred to the Web version of this article.)
In this study, we found that the color of model eggs can have an important effect on egg recognition in the Barn Swallow, a host of common cuckoos. They were more likely to reject white model eggs which are moderate mimetic to their own eggs, and blue model eggs (with shorter reflectance spectrum), which are highly non-mimetic to their own eggs, and were more likely to accept red model eggs (with longer reflectance spectrum), which are also highly non-mimetic to their own eggs. These results differ substantially from those of previous studies that hosts are more biased toward recognizing brown eggs and accepting blue eggs (Hanley et al., 2015, 2017, 2019; Abolins-Abols et al., 2019; but see Liu et al., 2024). The multiple rejection threshold model predicts that Barn Swallows should accept white model eggs and reject blue and red model eggs (Avilés, 2008; Cassey et al., 2008; Stoddard and Stevens, 2011; Stevens et al., 2013; Hauber et al., 2015; Manna et al., 2019). However, the Barn Swallows actually rejected more white and blue eggs than red eggs, which was not expected theoretically, as white eggs are more similar to their own eggs than blue and red eggs, at least seen from human eyes.
The single rejection threshold model predicts that hosts will accept blue/green and reject brown/red eggs (Hanley et al., 2017, 2019; Abolins-Abols et al., 2019); however, our experimental results generally show that Barn Swallows prefer to accept red eggs with longer wavelengths. In some cuckoo-host systems, hosts recognize foreign eggs but do not reject them (e.g. Moskát and Hauber, 2007; Antonov et al., 2009). However, in our case, Barn Swallows can easily peck the model eggs and reject them as the tested model egg are similar to their own eggs in egg size and mass (also see Yang et al., 2022a). Therefore, it is likely that Barn Swallows prefer to accept red eggs may also be due to the host's individual preference towards red color eggs rather than not recognizing red color eggs. The Barn Swallow is a suitable experimental subject for studying the host visual and cognitive mechanisms in brood parasitic birds, as they readily construct nests in village houses. It is precisely because Barn Swallows live alongside humans and utilize humans as a refuge from parasitism (Liang et al., 2013) that the parasitism rate of Barn Swallows is low, with a parasitism rate of 2.4% in Jilin and 0% in Hainan (Su et al., 2017). One possible reason for the lack of significant differences in egg recognition between Barn Swallows in different regions may be due to the low parasitism rate across all regions, and the differential impact of brood parasitism pressure on egg recognition might not be reflected. Alternatively, Barn Swallows may lack the parasitic pressure to recognize long wavelengths, may therefore not be able to reject red eggs, which needs to be confirmed by future investigations.
The results of foreign egg recognition by Barn Swallows partially confirmed to the single rejection threshold model, whereby blue eggs were recognized and ejected. Several studies examining the role of four separate photoreceptors in birds have found that perceived changes in ultraviolet and blue light predict host egg recognition behavior, whereas absolute color contrast differences do not (Cassey et al., 2008). Thus, color-specific cognitive changes may control host egg recognition behavior, and recognition responses may be adaptive if the host has an acquired or innate preference for rejecting or accepting the color of parasitic eggs (Hanley et al., 2017). As with other types of discrimination (Ghirlanda and Enquist, 2003; ten Cate and Rowe, 2007), host egg discrimination depends on a variety of factors, including sensory systems, cognitive abilities, coevolutionary history, and individual experience (Feeney et al., 2014). As brood parasitism pressure may vary, egg appearance (Lahti, 2005; Spottiswoode and Stevens, 2012) and response to parasitism (Soler et al., 2012) are likely to change over decades. Hence, if hosts preferentially reject brown and red eggs with longer wavelengths (e.g. Hanley et al., 2015, 2017), such as the Herring Gull (Larus argentatus) (Stratton and Dearborn, 2021), then it is likely that cuckoo egg-color variations with shorter wavelengths of blues and greens are more common than those actually found in nature (Fossøy et al., 2016). However, this needs to be verified through further investigation. In addition, a preference for red eggs has now been observed in certain birds (Stratton and Dearborn, 2021; Liu et al., 2024), whereas other host species do not exhibit a similar preference (Zhang et al., 2023), and further experimental verification involving more host species is still needed.
Our study suggests that the color of experimental eggs can influences egg rejection by the host. If a preference for certain visual stimuli (such as red eggs) by the host was the main selective force in the process of attaining egg acceptance, parasitic cuckoos would evolve the type of egg coloration that would be preferred and accepted by the host rather than that which is similar to the host's egg coloration. This would open new avenues to uncover the evolution of cuckoo egg mimicry, which needs to be verified in a larger number of host species.
Declaration of competing interests
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Ethics statement
The experiments comply with the current laws of China, where they were performed. Experimental procedures were in agreement with the Animal Research Ethics Committee of Hainan Provincial Education Centre for Ecology and Environment, Hainan Normal University (No. HNECEE-2012-001).
We would like to thank three reviewers for their constructive comments, which significantly improved this manuscript. We are grateful to Laikun Ma, Jianping Liu, Bo Zhou, Yuhan Zhang, Jinmei Liu and Kangning Luo for their help with the experiments and data collection.
Antonov, A., Stokke, B.G., Moksnes, A., Røskaft, E., 2009. Evidence for egg discrimination preceding failed rejection attempts in a small cuckoo host. Biol. Lett. 5, 169–171.
Abolins-Abols, M., Hanley, D., Moskát, C., Grim, T., Hauber, M.E., 2019. Anti-parasitic egg rejection by great reed warblers (Acrocephalus arundinaceus) tracks differences along an eggshell color gradient. Behav. Process. 166, 103902.
Avilés, J.M., 2008. Egg colour mimicry in the common cuckoo Cuculus canorus as revealed by modelling host retinal function. Proc. R. Soc. Lond. B Biol. Sci. 275, 2345–2352.
Campobello, D., Sealy, S.G., 2009. Avian brood parasitism in a Mediterranean region: hosts and habitat preferences of common cuckoos Cuculus canorus. Hous. Theor. Soc. 56, 399–400.
Canniff, L., Dainson, M., López, A.V., Hauber, M.E., Grim, T., Samaš, P., Hanley, D., 2018. Probing the limits of egg recognition using egg rejection experiments along phenotypic gradients. J. Visual. Exp. 138, e57512.
Cassey, P., Honza, M., Grim, T., Hauber, M.E., 2008. The modelling of avian visual perception predicts behavioural rejection responses to foreign egg colours. Biol. Lett. 4, 515–517.
Caves, E.M., Green, P.A., Zipple, M.N., Peters, S., Johnsen, S., Nowicki, S., 2018. Categorical perception of colour signals in a songbird. Nature 560, 365–367.
Feeney, W.E., Welbergen, J.A., Langmore, N.E., 2014. Advances in the study of coevolution between avian brood parasites and their hosts. Annu. Rev. Ecol. Evol. Syst. 45, 227–246.
Fossøy, F., Sorenson, M.D., Liang, W., Ekrem, T., Moksnes, A., Møller, A.P., et al., 2016. Ancient origin and maternal inheritance of blue cuckoo eggs. Nat. Commun. 7, 10272.
Ghirlanda, S., Enquist, M., 2003. A century of generalization. Anim. Behav. 66, 15–36.
Hanley, D., Grim, T., Cassey, P., Hauber, M.E., 2015. Not so colourful after all: eggshell pigments constrain avian eggshell colour space. Biol. Lett. 11, 20150087.
Hanley, D., Grim, T., Igic, B., Samaš, P., López, A.V., Shawkey, M.D., et al., 2017. Egg discrimination along a gradient of natural variation in eggshell coloration. Proc. R. Soc. Lond Biol. Sci. 284, 20162592.
Hanley, D., López, A.V., Fiorini, V.D., Reboreda, J.C., Grim, T., Hauber, M.E., 2019. Variation in multicomponent recognition cues alters egg rejection decisions: a test of the optimal acceptance threshold hypothesis. Philos. Trans. R. Soc. Lond. B Biol. Sci. 374, 20180195.
Hauber, M.E., Moskát, C., Bán, M., 2006. Experimental shift in hosts' acceptance threshold of inaccurate-mimic brood parasite eggs. Biol. Lett. 2, 177–180.
Hauber, M.E., Tong, L., Bán, M., Croston, R., Grim, T., Waterhouse, G.I., et al., 2015. The value of artificial stimuli in behavioral research: making the case for egg rejection studies in avian brood parasitism. Ethology 121, 521–528.
Igic, B., Cassey, P., Grim, T., Greenwood, D.R., Moskát, C., Rutila, J., et al., 2012. A shared chemical basis of avian host-parasite egg colour mimicry. Proc. R. Soc. Lond. B Biol. Sci. 279, 1068–1076.
Lahti, D.C., 2005. Evolution of bird eggs in the absence of cuckoo parasitism. Proc. Natl. Acad. Sci. U.S.A. 102, 18057–18062.
Lahti, D.C., 2006. Persistence of egg recognition in the absence of cuckoo brood parasitism: pattern and mechanism. Evolution 60, 157–168.
Liang, W., Yang, C., Wang, L., Møller, A.P., 2013. Avoiding parasitism by breeding indoors: cuckoo parasitism of hirundines and rejection of eggs. Behav. Ecol. Sociobiol. 67, 913–918.
Liu, J., Wang, X., Liang, W., 2024. Attractive egg color? Red eggs facilitate egg acceptance in a cavity-nesting host. Curr. Zool. 70, 112–114.
Manna, T., Moskát, C., Hauber, M.E., 2017. Cognitive decision rules for egg rejection. In: Soler, M. (Ed.), Avian Brood, Parasitism: Behaviour, Ecology, Evolution and Coevolution. Springer, Cham, pp. 437–448.
Manna, T.J., Moskát, C., Tong, L., Bán, M., Aidala, Z., Low, J., et al., 2019. Multiple parasitism reduces egg rejection in the host (Acrocephalus arundinaceus) of a mimetic avian brood parasite (Cuculus canorus). J. Comp. Psychol. 133, 351–358.
Moskát, C., Hauber, M.E., 2007. Conflict between egg recognition and egg rejection decisions in common cuckoo (Cuculus canorus) hosts. Anim. Cognit. 10, 377–386.
Reeve, H.K., 1989. The evolution of conspecific acceptance thresholds. Am. Nat. 133, 407–435.
Rothstein, S.I., 1990. A model system for coevolution: avian brood parasitism. Annu. Rev. Ecol. Evol. Syst. 21, 481–508.
Scordato, E.S., Safran, R.J., 2014. Geographic variation in sexual selection and implications for speciation in the barn swallow. Avian Res. 5, 8.
Soler, M., Martín-Vivaldi, M., Fernández-Morante, J., 2012. Conditional response by hosts to parasitic eggs: the extreme case of the rufous-tailed scrub robin. Anim. Behav. 84, 421–426.
Soler, M., 2014. Long-term coevolution between avian brood parasites and their hosts. Biol. Rev. 89, 688–704.
Spottiswoode, C.N., Stevens, M., 2012. Host-parasite arms races and rapid changes in bird egg appearance. Am. Nat. 179, 633–648.
Stevens, M., Troscianko, J., Spottiswoode, C.N., 2013. Repeated targeting of the same hosts by a brood parasite compromises host egg rejection. Nat. Commun. 4, 2475.
Stoddard, M.C., Stevens, M., 2011. Avian vision and the evolution of egg color mimicry in the common cuckoo. Evolution 65, 2004–2013.
Stratton, B.J., Dearborn, D.C., 2021. Nest sanitation behavior does not increase the likelihood of parasitic egg rejection in herring gulls. Curr. Zool. 67, 675–682.
Su, T., Huo, J., Yang, C., Liang, W., 2017. Brood parasitism on barn swallow (Hirundo rustica) populations in China by common cuckoo (Cuculus canorus). Chin. J. Zool. 52, 338–341.
Šulc, M., Hughes, A.E., Mari, L., Troscianko, J., Tomášek, O., Albrecht, T., et al., 2022. Nest sanitation as an efective defence against brood parasitism. Anim. Cognit. 25, 991–1002.
Tian, L., Liu, Y., Wu, Y., Feng, Z., Hu, D., Zhang, Z., 2024. Migration pattern of a population of barn swallows (Hirundo rustica) breeding in East Asian tropical region. Avian Res. 15, 100192.
ten Cate, C., Rowe, C., 2007. Biases in signal evolution: learning makes a difference. Trends Ecol. Evol. 22, 380–387.
Turner, A., 2006. The Barn Swallow. T & AD Poyser, London.
Yang, C., Chen, M., Wang, L., Liang, W., Møller, A.P., 2015a. Nest sanitation elicits egg discrimination in cuckoo hosts. Anim. Cognit. 18, 1373–1377.
Yang, C., Liang, W., Møller, A.P., 2019. Using 3D modelling and printing to study avian cognition from different geometric dimensions. R. Soc. Open Sci. 6, 181938.
Yang, C., Møller, A.P., Liang, W., 2022b. Light matters: nest illumination alters egg rejection behavior in a cavity-nesting bird. Avian Res 13, 100016.
Yang, C., Wang, L., Liang, W., Møller, A.P., 2015b. Nest sanitation behavior in hirundines as a pre-adaptation to egg rejection to counter brood parasitism. Anim. Cognit. 18, 355–360.
Yang, Q., Chen, X., Zhang, Z., Han, J., Wu, N., Yang, C., 2022a. Egg rejection and nest sanitation in an island population of barn swallows (Hirundo rustica): probability, response latency, and sex effects. Animals 12, 3027.
Zhang, Y., Zhong, G., Wan, G., Wang, L., Liang, W., 2023. Brood parasitism and egg recognition in three bunting hosts of the cuckoos. Ecol. Evol. 13, e10659.
Zink, R.M., Pavlova, A., Rohwer, S., Drovetski, S.V., 2006. Barn swallows before barns: population histories and intercontinental colonization. Proc. R. Soc. Lond. B Biol. Sci. 273, 1245–1251.
Zipple, M.N., Caves, E.M., Green, P.A., Peters, S., Johnsen, S., Nowicki, S., 2019. Categorical colour perception occurs in both signalling and non-signalling colour ranges in a songbird. Proc. R. Soc. Lond. B Biol. Sci. 286, 524.


 DownLoad:
DownLoad:





 Email Alerts
Email Alerts RSS Feeds
RSS Feeds