Csaba MOSKÁT, Fugo TAKASU, A. Roman MUÑOZ, Hiroshi NAKAMURA, Miklós BÁN, Zoltán BARTA. 2012: Cuckoo parasitism on two closely-related Acrocephalus warblers in distant areas: a case of parallel coevolution?. Avian Research, 3(4): 320-329. DOI: 10.5122/cbirds.2012.0038
Citation:
Csaba MOSKÁT, Fugo TAKASU, A. Roman MUÑOZ, Hiroshi NAKAMURA, Miklós BÁN, Zoltán BARTA. 2012: Cuckoo parasitism on two closely-related Acrocephalus warblers in distant areas: a case of parallel coevolution?. Avian Research, 3(4): 320-329. DOI: 10.5122/cbirds.2012.0038
Csaba MOSKÁT, Fugo TAKASU, A. Roman MUÑOZ, Hiroshi NAKAMURA, Miklós BÁN, Zoltán BARTA. 2012: Cuckoo parasitism on two closely-related Acrocephalus warblers in distant areas: a case of parallel coevolution?. Avian Research, 3(4): 320-329. DOI: 10.5122/cbirds.2012.0038
Citation:
Csaba MOSKÁT, Fugo TAKASU, A. Roman MUÑOZ, Hiroshi NAKAMURA, Miklós BÁN, Zoltán BARTA. 2012: Cuckoo parasitism on two closely-related Acrocephalus warblers in distant areas: a case of parallel coevolution?. Avian Research, 3(4): 320-329. DOI: 10.5122/cbirds.2012.0038
MTA-ELTE-MTM Ecology Research Group, Hungarian Academy of Sciences, c/o Biological Institute, Eötvös Lóránd University, Pázmány Péter sétány 1/C., H-1117 Budapest, Hungary and Hungarian Natural History Museum, Baross u. 13., H-1088 Budapest, Hungary
2.
Department of Information and Computer Sciences, Nara Women's University, Kita-Uoya Nishimachi, Nara 630-8506, Japan
3.
Fundación Migres, N-340 Km. 96, Algeciras, E-11390, Cádiz, Spain
4.
Department of Biology, Faculty of Education, Shinshu University, Nishinagano, Nagano 380-8544, Japan
5.
MTA-DE "Lendület" Behavioural Ecology Research Group, Department of Evolutionary Zoology, University of Debrecen, H-4010 Debrecen, Hungary
Common Cuckoos (Cuculus canorus) parasitize nests of small passerines. The Cuckoo chicks cause the death of their nest-mates when evicting eggs or nestlings from the nests; consequently, hosts suffer from a high loss of reproduction. Host adaptations against parasitism, e.g., by egg discrimination behavior, and cuckoo counter-adaptations to hosts, e.g., by mimetic eggs, are often regarded as a result of the arms race between the two interacting species. In Hungary Great Reed Warblers (Acrocephalus arundinaceus) are the main hosts of cuckoos, suffering from heavy parasitism (ca. 40-65%). The Oriental Reed Warbler (A. orientalis), formerly a subspecies of the Great Reed Warbler (A. a. orientalis), is also a highly parasitized host in Japan (25-40%). We compared main characteristics of Cuckoo parasitism in these two distant areas from the Western and Eastern Palearctic by comparing cuckoo egg mimicry. We measured color characteristics of host and parasitic eggs by spectrophotometer. Visual modeling revealed lower chromatic distances between Cuckoo and host eggs in Hungary than in Japan, but high variation both in host and Cuckoo eggs may cause matching problems in Hungary. Achromatic (brightness) difference between host and Cuckoo eggs were lower in Japan than in Hungary, and it proved to be the most important factor affecting egg rejection. Hosts rejected Cuckoo eggs at similar frequencies (37% and 35% in Hungary and Japan, respectively). Host adaptation, i.e., egg rejection behavior, seems to be preceding Cuckoo counter-adaptations to hosts in Japan. We suggest that the Cuckoo-Great/Oriental Reed Warbler relationships developed in alternative ways in Japan and Hungary, and they represent different stages of their arms race.
The brood parasitic Common Cuckoos (hereafter "Cuckoo"; Cuculus canorus) lay eggs into nests of other bird species, and the Cuckoo chick evicts all nest contents, eggs or nestlings, after hatching, typically causing the failure of all host offspring. This represents a high cost of parasitism for the hosts (Davies, 2000; Kilner, 2006). However, future prospects of the cuckoo eggs depend on host egg recognition and their decision on egg discrimination. Hosts may possess the ability to recognize and reject the parasite egg (Davies and Brooke, 1989; Moksnes et al., 1991; Hauber and Sherman, 2001), so natural selection favors the development of mimetic foreign eggs (Davies, 2000). Mimicry of the parasites' eggs in relation to host eggs may reach high levels in some populations (Moksnes and Røskaft, 1995; Igic et al., 2012).
In Japan, Common Cuckoos also parasitize a closelyrelated Acrocephalus species, the Oriental Reed Warbler (Acrocephalus orientalis), which previously was not separated from the Great Reed Warbler, but regarded as one of its subspecies (A. arundinaceus orientalis). Oriental Reed Warblers are also parasitized in a relatively high frequency (8–22%; Lotem et al., 1995), but lower than Great Reed Warblers in Hungary (see above). The Japanese population of Oriental Reed Warblers at Nagano City also showed antiparasite defenses against Cuckoo eggs, where different studies revealed 62% (Lotem et al., 1995) or 45% rejection rates towards Cuckoo eggs (Nakamura et al., 1998).
As both the Japanese and the Hungarian populations of the above-mentioned Acrocephalus hosts show moderately developed antiparasite defenses, it may be a reflection of that both the host and brood parasite populations have been in contact with each other. Although the two populations of the two studied warblers may be different in their evolutionary history, including their coevolution with the Cuckoo, this case offers a unique situation for the comparison of egg mimicry and antiparasite defense in two distant areas. Hostbrood parasite coevolution might take several alternative directions regarding egg appearances in time and space (Dawkins and Krebs, 1979; Rothstein, 1990), while still maintaining the adaptive value of egg coloration (Underwood and Sealy, 2002; Kilner, 2006). Although brood parasitism is a dominant selective agent forcing the development of Cuckoo egg characteristics (Stokke et al., 2002; Kilner, 2006), we have no previous knowledge if hosts and brood parasites follow the same scenario in their coevolution at two distant sites. However, modeling coevolution has revealed several alternative potential pathways for Cuckoos and their hosts (Takasu, 1998, 2003; Barabás et al., 2004; Røskaft et al., 2006; Takasu and Moskát, 2011).
In this paper, we compared main characteristics of Cuckoo parasitism on two relatively large-sized Acrocephalus species in Japan and Hungary. We analyzed spectrophotometric data by visual models taking into account the sensitivity to different wavelengths of the four cone types in the avian retina (Endler and Milke, 2005). This method let us incorporate the effects of avian perceptual properties as it has recently been applied and recommended for the comparison of color attributes of Cuckoo and host eggs (e.g., Cassey et al., 2008; Avilés, 2008; Langmore et al., 2009; Antonov et al., 2010; Stoddard and Stevens, 2010; Vikan et al., 2011). Following the idea of the arms race hypothesis (Dawkins and Krebs, 1979; Davies, 2000; Krüger, 2007) applied for the coevolution between the Cuckoo and its hosts, we predicted that Cuckoos developed adaptations to host rejection by developing mimetic eggs in both areas. We also examined host responses to Cuckoo parasitism in the two distant areas, and predicted higher similarities between accepted eggs and lower similarities between eggs which were rejected. Finally, we predicted that the Cuckoo egg mimicry and host egg rejection behavior are similar in the two areas, indicating a similar level of coevolutionary adaptations.
Methods
The study was carried out at two sites: (1) Nobeyama highlands (35°57'N, 138°28'E), ca. 120 km south of Nagano city, central Japan. In this area Oriental Reed Warblers breed in small patches of reeds (Phragmites australis and Miscanthus sacchariflorus) along small (1–2 m wide) streams, and reed patches are surrounded by wooded vegetation composed primarily of maple trees (Acer ginnala) and willow bushes (Salix integra). The study was conducted during May–July 2004. (For more details of the habitat, see Muñoz et al., 2007). (2) In the surroundings of Apaj (47°07'N, 19°06'E), ca. 50 km south of Budapest in the Hungarian Great Plains, central Hungary. Here Great Reed Warblers breed in 2–4 m wide stripes of reed (Phragmites australis) growing along both sides of small (5–10 m wide) channels. Lines of trees (hybrid poplars, Populus alba, Salix cinerea, Elaeagnus angustifolia) and bushes typically follow the channels, although treeless sections also occur. (For more details of the habitat, see Moskát and Honza, 2000, 2002). Only those nests containing one Cuckoo egg and at least one host egg, and if the hosts' response to parasitism was known, were used for analyses.
We systematically searched for nests in parts of the study area ca. twice per week. Nests in the egg laying stage were controlled daily. Nests parasitized by a Cuckoo were controlled for six consecutive days to observe host reactions, and used to categorize host responses to parasitism. This 6-day (or 5-day) long period for monitoring hosts' reactions is typically used in studies of natural Cuckoo parasitism (e.g., Lotem et al., 1995; Moskát and Honza, 2002) and experiments with model Cuckoo eggs (e.g., Moksnes et al., 1991; Bártol et al., 2002). Host responses were categorized as acceptance or rejection of the Cuckoo egg. The term rejections included egg ejections and nest desertions (see more details on the method of nest monitoring in Moskát and Honza, 2002).
Eggs were photographed using digital cameras (Olympus E-20 and Nikon CoolPix 995) on Kodak grey card with Kodak control color patches. We measured reflectance spectra (300–700 nm) from the eggshells by spectrophotometers type Ocean Optics USB 2000, which interval covers the range of the vision of small passerines (e.g., Bennett and Cuthill, 1994; Endler and Milke, 2005). All eggs were measured in a clutch, although not all clutches were measured by spectrophotometer, so sample sizes slightly differ for host responses and egg characteristics. We measured reflectances at eight points on the eggshell, i.e., two measurements at four zones of the eggs, dividing the eggshell surface into four equally long parts along the long axis (see for more details on measuring reflectances from eggs in Takasu et al., 2009). A conceptually similar method, when the eggshell surface is divided into segments along the long axis, was developed by Cherry and Bennett (2001).
For the evaluation of chromatic difference (∆TC) among Cuckoo and host eggs we applied the perceptual models developed for avian vision (Kelber et al., 2003; Endler and Milke, 2005). These models provide a useful way for the quantification of color difference between eggs as birds can see, with respects to the spectral sensitivities of the avian single cone photoreceptors in the retina (Vorobyev and Osorio, 1998). In birds the four single photoreceptors are sensitive to certain parts of the whole spectra within 300 and 700 nm, called ultraviolet, short-wavelength, medium-wavelength and long-wavelength (Hart et al., 2000). In comparison of color differences between Common Cuckoo and host eggs these models have already been useful (e.g., Avilés, 2008; Cassey et al., 2008; Antonov et al., 2010; Vikan et al., 2011), and also for other brood parasite-host relationships (Langmore et al., 2009; Spottiswoode and Stevens, 2010). We used the "woodland canopy-filtered green light" (also called "green leaves"; Endler, 1993) and "open nest" (Avilés, 2008) ambient light conditions for ambient irradiance in the perceptual models, as Acrocephalus warblers typically breed in open nests in the reeds, in the shade of long reed leaves. We also calculated achromatic contrast (∆TQ) expressing brightness differences between Cuckoo and host eggs. Chromatic and achromatic differences are expressed in just noticeable difference thresholds (JNDs), where a value above the JND = 1 threshold suggests that the cuckoo egg can be discriminated from the host eggs, but a value below this threshold suggests that the parasitic egg is undistinguishable (Osorio and Vorobyev, 1996, see for more details in Avilés, 2008 and Igic et al., 2012). For visual modeling we used Avicol version 4 (Gomez, 2010).
In the present study we focused on the importance of chromatic and achromatic components of eggshell characteristics in coevolution between brood parasites and their hosts, but spottiness may also have some relevance (Polaciková and Grim, 2010). However, the fiber optic probe of our spectrophotometer only allows samples from larger spots, only > 2 mm in diameter, which is not suitable for speckles or lines. Experimental studies on the Hungarian Great Reed Warbler population has shown that small spots may be important in egg recognition (Moskát et al., 2008a), but the role of larger spots is not so clear (Moskát et al., 2008c).
We used generalized linear models to test the dependent variable (acceptance versus rejection), involving intercept, country as independent fixed factor, chromatic difference (∆TC), and achromatic difference (∆TQ) as covariates. We also involved the interaction terms chromatic difference×country and achromatic difference ×country. Data were analyzed with SPSS version 17.0 (SPSS, Inc.). We applied the Levene's test for testing homogeneity of variances prior to parametric tests.
Results
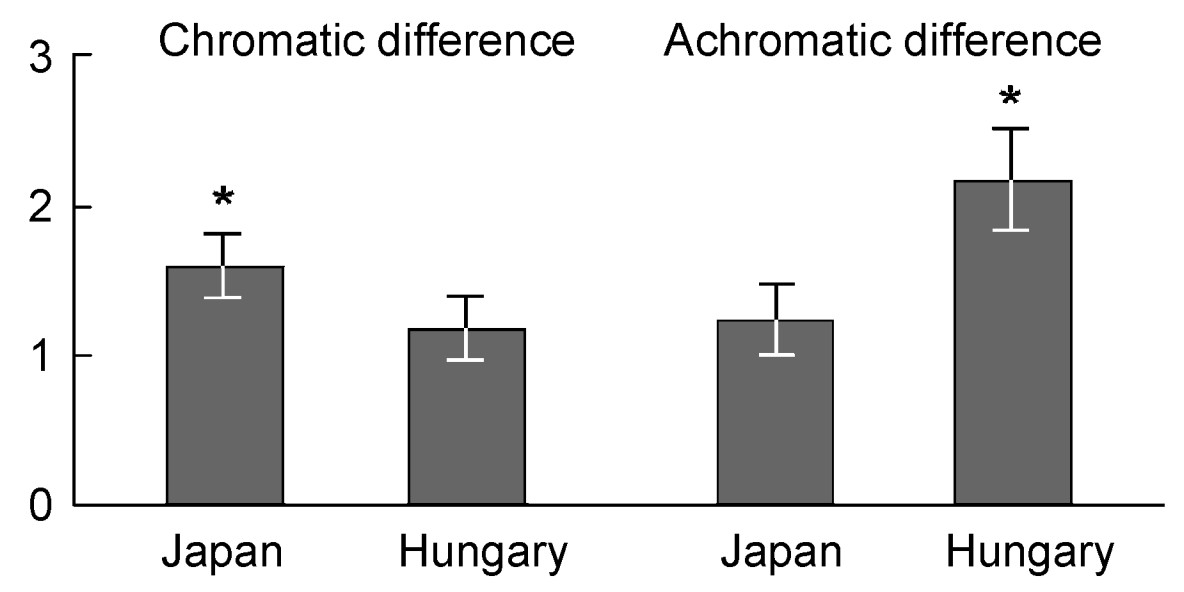
Some of the Cuckoo eggs found in Hungary showed a good mimicry to host eggs, while in general mimicry was lower in Japan (Fig. 1). Although the main characteristics of host eggs were similar in Japan and Hungary, Cuckoos in Japan often showed red markings, sometimes with the special line pattern (Fig. 1). Visual modeling on spectrophotometer measurements revealed that chromatic difference between Cuckoo and host eggs was lower in Hungary than in Japan, whereas the achromatic difference showed the opposite trend (Fig. 2; open nest ambient light conditions, Mann-Whitney U-tests were significant for both the chromatic and achromatic cases: U = 100.00, p = 0.049). The values of chromatic difference did not differ significantly from the just noticeable difference threshold (JND = 1) (ones-sample t-test, t17 = 0.910, p = 0.375) in Hungary, which means that Cuckoo eggs are more difficult to recognize in Hungary than in Japan, where chromatic difference exceeded the threshold significantly (t17 = 2.948, p = 0.009). However, the achromatic difference between parasite and host eggs did not differ from the threshold in Japan (two-sample t-test, t17 = 0.867, p = 0.398), but it was significantly different in Hungary (t17 = 3.244, p = 0.005). We found similar results when the green ambient light conditions was used for calculation (Hungary: chromatic contrast: t17 = 0.505, p = 0.620, achromatic contrast: t17 = 3.246, p = 0.005; Japan: chromatic contrast: t17 = 2.461, p = 0.025, achromatic contrast: t17 = 0.887, p = 0.388).
Figure
1.
Types of egg morphs of Common Cuckoos (Cuculus canorus) and their two closely-related Acrocephalus hosts from Hungary (host: Great Reed Warbler, A. arundinaceus) and Japan (host: Oriental Reed Warbler, A. orientalis)
Figure
2.
Mean perceived differences (±S.E.) for the chromatic and achromatic components of color matching between the eggs of Common Cuckoos and their two closely-related Acrocephalus hosts, i.e., the Oriental Reed Warbler in Japan and Great Reed Warbler in Hungary. Asterisks indicate significant difference from the just noticeable difference threshold, JND = 1, at the p ≤ 0.05 level, obtained by onesample t-test.
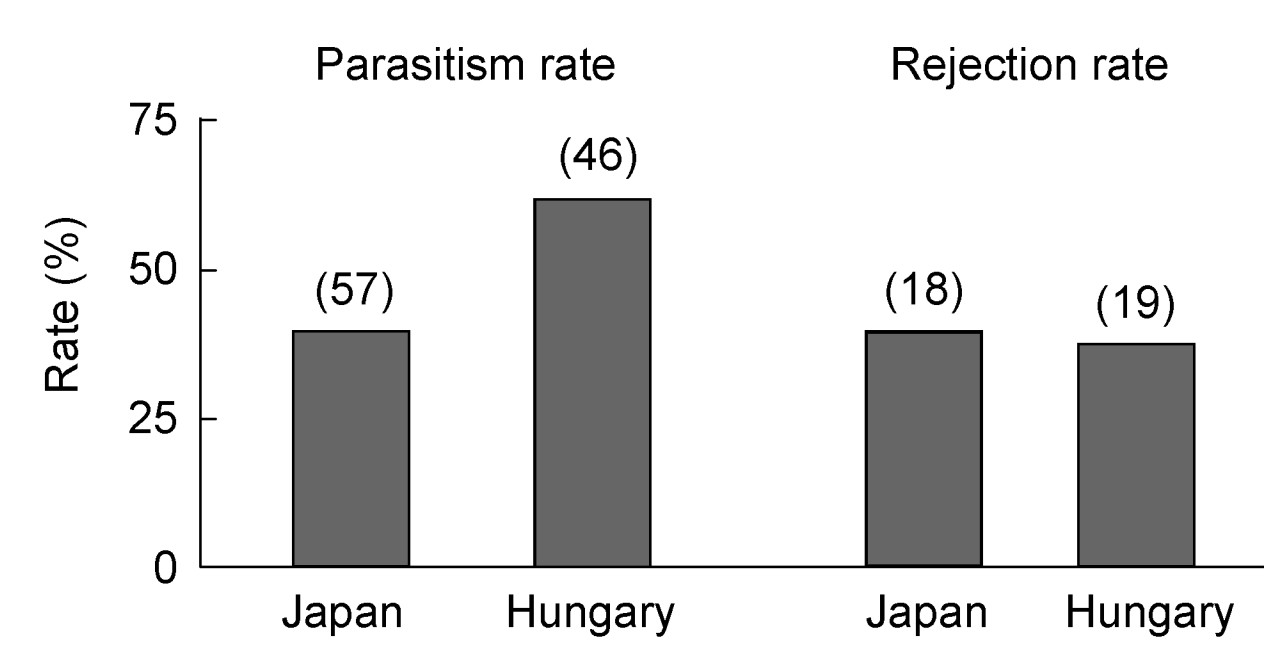
We observed host reactions toward Cuckoo eggs in 37 nests. Rejection rates against single Cuckoo eggs were similar at the two sites (7/19 in Hungary and 7/18 in Japan; Fig. 3; χ12 = 0.02, p = 0.898). Parasitism rate was higher in Hungary than in Japan (28/46 nests, i.e., 61% parasitism rate in Hungary and 22/57 nests, i.e., 39% in Japan in 2004), however, the difference was not statistically significant (χ12 = 1.73; p = 0.188).
Figure
3.
Parasitism rates and rejection rates of Common Cuckoo eggs in two closely-related Acrocephalus species at distant areas, i.e., the Oriental Reed Warbler in Japan and Great Reed Warbler in Hungary
Generalized linear models on egg characteristics of the two Acrocephalus species revealed no direct effect of chromatic and achromatic difference between host and parasite eggs on the rejection of Cuckoo eggs. However, we revealed significant effects of country and the term of interaction between country and achromatic distance (∆TQ) in the generalized linear model, while achromatic distance without the interaction term proved not to be significant (country: χ2 = 3.90, p = 0.048; ∆TC: χ2 = 0.24, p = 0.628; ∆TQ: χ2 = 3.31, p = 0.069; country×∆TC: χ2 = 0.41, p = 0.524; country×∆TQ: χ2 = 5.90, p = 0.015; intercept: χ2 = 0.04, p = 0.853).
Discussion
Cuckoo egg mimicry
Regarding eggshell colors, our analyses revealed a higher similarity of Cuckoo eggs with respect to host eggs in Hungary than in Japan. It is an important indicator of coevolutionary adaptations, as mimicry of Cuckoo eggs is expected to increase over time during Cuckoo parasitism (see review by Davies, 2000). Although exact time when Cuckoo parasitism started is not known, neither for the Great Reed Warbler in Hungary, nor the Oriental Reed Warbler in Japan, circumstantial evidence suggests that the history of these coevolutionary interactions reflects a longer history in the Western Palearctic than in the Eastern Palearctic. Recent Cuckoo parasitism is estimated for more than 110 years in Hungary, since parasitized Great Reed Warbler clutches with Cuckoo eggs were already collected ca. 110 years ago and stored in museums (C. Moskát unpubl.). Cuckoo parasitism started ca. 50 years ago in the Nobeyama district, but probably it started earlier in other parts of Japan (Nakamura, 1990). In Hungary, Cuckoos have developed almost perfect mimicry regarding background and spot colors and spottiness of eggs, but not in egg shape (Bán et al., 2011). On the one hand, high interclutch variation of hosts makes it more difficult for the Cuckoo to match the host eggs even when Cuckoo eggs show perfect mimesis to one type of Great Reed Warbler eggs (Moskát and Honza, 2002). On the other hand Cuckoos selecting actively for proper nests may increase their success of matching (Cherry et al., 2007b; see also Avilés et al., 2006).
Cuckoo eggs in the Hungarian study area are found to be polymorphic (Moskát and Honza, 2002), therefore, even cuckoo eggs showing poor mimesis to host eggs can be found in the area. For example, one poorly mimetic egg type, the so-called Garden Warbler (Sylvia borin)-type Cuckoo egg was also found in the study area, which seems to be a generalist Cuckoo egg-type in central Europe (Lovászi and Moskát, 2004). This egg type has also been reported from the Czech Republic (Edvardsen et al., 2001; Honza et al., 2001). Interestingly, the Garden Warbler-type Cuckoo egg morph found in Hungary had a high similarity with Garden Warbler eggs from Germany, but this egg morph is different from Garden Warbler eggs in Hungary (C. Moskát, unpubl.). In the 1920s a system of flood relief and irrigation channels was established in Hungary, offering a preferable habitat with narrow reed beds along both sides of the channels. This habitat offered the open water surface-reed edge for birds, which is favored by Great Reed Warblers (Báldi and Kisbenedek, 2000). Although in the Hungarian Great Plains wetland habitats dominated originally, they were lost gradually. However, extensive work to control rivers' flood during the second half of the 19th century made numerous dead branches, especially suitable habitats for the Great Reed Warbler (Molnár, 1944). We therefore suppose that Cuckoos started to parasitize this warbler extensively in the dead branches, and, later, in the channels depending on the availability of trees used for vantage points by nest searching Cuckoos (Moskát and Honza, 2000). The possible origin of recent Cuckoo parasitism on the Great Reed Warbler was the parasitism on several Acrocephalus species, including the Great Reed Warbler, Reed Warbler (A. scirpaceus), Sedge Warbler (A. schoenobaenus) and Marsh Warbler (A. palustris), because parasitized clutches of these species can be found from the first half of the 20th century in Hungarian bird egg collections (C. Moskát, unpubl.). However, recent parasitism on these three smaller Acrocephalus species seems to be scarce (C. Moskát, unpubl.).
In Japan the different Cuckoo gentes are less separated than in Europe (Higuchi, 1998; Nakamura et al., 1998). The Bunting-type Cuckoo eggs seem to be the ancient form, and probably Cuckoos of several host species radiated from this gens (c.f. Nakamura et al., 1998; Takasu et al., 2009). Although the Siberian Meadow Bunting (Emberiza cioides) used to be the most Common Cuckoo host in the first half of the 20th century (Nakamura et al., 1998), it is presently almost abandoned by Cuckoos. This host species developed high level egg discrimination ability, and, consequently, Cuckoos expanded the number of their hosts. Many Cuckoo eggs exhibit small lines as pigments on the eggshell (Nakamura et al., 1998; Takasu et al., 2009), which is not typical for European Cuckoos. In our study area at Nobeyama, in central Japan, four host species are frequently parasitized by Cuckoos: Black-faced Buntings (E. spodocephala), Bull-headed Shrikes (Lanius bucephalus), Azure-winged Magpies (Cyanopica cyana) and Oriental Reed Warblers. Cuckoo eggs parasitizing these four host species seem to be similar, but some early separation was revealed by a recent spectrophotometer study (Takasu et al., 2009). The bunting-type Cuckoo egg morph matches well the eggs of Bull-headed Shrikes, but it is non-mimetic for the Azure-winged Magpie and Oriental Reed Warbler. So the Japan Cuckoo-host relationship is a sympatric multihost system, as Cuckoos parasitize several host species at the same location. Although these hosts prefer different habitats, the imperfect isolation of these host populations from each other slows down adaptational mechanisms (sensu Møller et al., 2011). The presence of lines as egg markings on Cuckoo eggs in Japan is unique when compared either to Cuckoo eggs in Hungary, or any of the studied Acrocephalus hosts (Fig. 1). This special marking on Cuckoo eggs may also reduce mimicry between Oriental Reed Warbler and Cuckoo eggs. Contrary, in the Hungarian Great Plane Cuckoos parasitize only one dominant host species, the Great Reed Warbler. There is another frequently used host species in Hungary, the Robin (Erithacus rubecula), which shows a strong separation from Great Reed Warblers by habitat, as Robins occur in the woodlands of hilly areas (Fuisz and de Kort, 2007).
Predictors of hosts' egg rejection behavior
Rejection rates in the two Acrocephalus Warbler populations proved to be highly similar, despite the differences in Cuckoo egg mimicry. This finding suggests that coevolutionary mechanisms, i.e., hosts' antiparasite defences, and Cuckoos' counter-adaptations to hosts by mimetic eggs may have different speed. Thus coevolutionary interactions are probably less synchronized than predicted by the arms race model (Dawkins and Krebs 1979; Davies 2000; Krüger 2007). This might be due to hosts' immigration from nearby, highly productive, probably unparasitized or less parasitized source populations in a metapopulation structure (Barabás et al., 2004). Immigration of hosts with less developed antiparasite defense might result in reduced egg discrimination ability in the Hungarian study area. This process might explain the similar level of egg rejection in Japan and Hungary, even though, the Hungarian Cuckoo-Great Reed Warbler system is more developed, as indicated by the higher level of mimesis of Cuckoo eggs here.
A previous study on the Oriental Reed Warbler in Nagano district, Japan (ca. 120 km away from our study www.chinesebirds.net site in Nobeyama district) revealed an even higher rejection rate than our study revealed (62% vs 39%) towards natural Cuckoo eggs between 1989 and 1991 (Lotem et al., 1995). The population in Nagano district started to be parasitized only 30 years ago, so it is likely that some parts of egg discrimination is the result of a former interaction with the Cuckoo, or due to gene flow from other areas (Lotem et al., 1995).
Parallel coevolution: the same scenario or alternative pathways?
Our results revealed similar efficiency in egg rejection behavior of the two Acrocephalus studied hosts, which can be regarded as one of the most important indicators of hosts' evolutionary adaptation to brood parasitism. On the other hand, we also revealed better mimicry of Cuckoo eggs to host eggs in Hungary than in Japan. The origin of Cuckoo parasitism on the Great Reed Warbler in Hungary differs from that of the Oriental Reed Warbler in Japan. While Cuckoos might have radiated from the Bunting Cuckoo-gens in Japan, in Hungary the Great Reed Warbler-Cuckoo gens probably originated from the Acrocephalus group. It is a surprising result that Japanese Cuckoos show low level mimicry to Oriental Reed Warbler eggs, although this warbler species possess a moderate egg rejection ability. Theoretically, in such type of systems better parasite egg mimicry is expected, as simulation models have revealed (Takasu, 2003). There are two possible explanations for this interesting result. At one hand there is a multihost system in central Japan, which might slow down separation of Cuckoos into distinct gentes (c.f. Higuchi, 1998; Nakamura et al., 1998). Takasu (2003) showed by simulation models, if the host population is polymorphic regarding egg morphs, this might prevent Cuckoos to develop a better mimicry.
Host aggression against the Cuckoo may also be evolved during sympatry with the brood parasite, and typically coevolves with egg rejection ability (Røskaft et al., 2002). As hosts breed in similar habitats in our two study sites, we think that host defense plays the most important role in Cuckoo-host coevolutions in these cases. In Hungary Great Reed Warblers show high aggression against the Cuckoo (85% attack, n = 40; Bártol et al., 2002), but Oriental Reed Warblers attacked Cuckoos at a lower frequency (38%; Andou et al., 2005). However, sample size in the Japanese study was low (n = 8).
We conclude that despite of the dissimilar features of Cuckoo-Warbler coevolution in Japan and Hungary, including possible different origins of parasitism and levels of parasitic pressure, Cuckoos showed adaptations to host eggs at varying degrees of egg mimicry, and hosts developed egg discrimination behavior against Cuckoo eggs. Hosts' egg discrimination proved to be very similar in the two host-brood parasite systems, although the pattern of egg mimicry is clearly different. Our results suggest that Cuckoos and Warblers followed different pathways of coevolution in the two distant areas, and they seem to be at different stages in the arms race.
Acknowledgments
The authors are thankful to Tibor Kisbenedek, László Bagó, István Bártol and Kinga Iglói for their assistance in the field work. For discussions we thank Maria Altamirano, Michael I. Cherry, Marcel Honza, Rita Hargitai, Branislav Igic and many other colleagues. We thank two anonymous reviewers and the guest editors for helpful comments on our manuscript. The study was supported by the bilateral project of the Japan Society for the Promotion of Science (JSPS) and the Hungarian Academy of Sciences to F.T. and C.M. 2003-2005, and by the Hungarian Scientific Research Fund (OTKA; grant No. T35015, 48397 and 83217) to C.M. The Duna-Ipoly and Kiskunság National Parks provided permissions for research.
Andou D, Nakamura H, Oomori S, Higuchi H. 2005. Characteristics of brood parasitism by Common Cuckoos on Azurewinged Magpies, as illustrated by video recordings. Avian Sci, 4:43–48.
Antonov A, Stokke BG, Vikan JR, Fossøy F, Ranke PS, Røskaft E, Moksnes A, Møller AP, Shykoff JA. 2010. Egg phenotype differentiation in sympatric cuckoo Cuculus canorus gentes. J Evol Biol, 23:1170–1182.
Avilés JM, Soler JJ, Soler M, Møller AP. 2006. Rapid increase in cuckoo egg matching in a recently parasitized reed warbler population. J Evol Biol, 19:1901–1910.
Avilés JM. 2008. Egg colour mimicry in the common cuckoo Cuculus canorus as revealed by modelling host retinal function. Proc R Soc B, 275:2345–2352.
Báldi A, Kisbenedek T. 2000. Bird species numbers in an archipelago of reeds at Lake Velence, Hungary. Global Ecol Biogeogr, 9:451–461.
Bán M, Barta Z, Muñoz RA, Takasu F, Nakamura H, Moskát C. 2011. The analysis of common cuckoo's egg shape in relation to its hosts' in two geographically distant areas. J Zool, 284:77–83.
Barabás L, Gilicze B, Takasu F, Moskát C. 2004. Survival and antiparasite defense in a host metapopulation under heavy brood parasitism: a source-sink dynamic model. J Ethol, 22:143–151.
Bártol I, Karcza Z, Moskát C, Røskaft E, Kisbenedek T. 2002. Responses of great reed warblers Acrocephalus arundinaceus to experimental brood parasitism: the effects of a cuckoo Cuculus canorus dummy and egg mimicry. J Avian Biol, 33:420–425.
Begum S, Moksnes A, Røskaft E, Stokke BG. 2011. Interactions between the Asian koel (Eudynamys scolopacea) and its hosts. Behaviour, 148:325–340.
Bennett ATD, Cuthill IC. 1994. Ultraviolet vision in birds: what is its function? Vision Res, 34:1471–1478.
Brooke M de L, Davies NB. 1988. Egg mimicry by Cuckoos Cuculus canorus in relation to discrimination by hosts. Nature, 335:630–632.
Cassey P, Honza M, Grim T, Hauber ME. 2008. The modelling of avian visual perception predicts behavioural rejection responses to foreign egg colours. Biol Lett, 45:15–517.
Cherry MI, Bennett ATD, Moskát C. 2007a. Host intra-clutch variation, cuckoo egg matching and egg rejection by great reed warblers. Naturwissenschaften, 94:441–447.
Cherry MI, Bennett ATD, Moskát C. 2007b. Do cuckoos choose nests of great reed warblers on the basis of host egg appearance? J Evol Biol, 20:1218–1222.
Cherry MI, Bennett ATD. 2001. Egg colour matching in an African cuckoo as revealed by ultraviolet-visible reflectance spectrophotometry. Proc R Soc B, 268:565–571.
Davies NB, Brooke M de L. 1988. Cuckoos versus reed warblers: adaptations and counteradaptations. Anim Behav, 36:262–284.
Davies NB, Brooke M de L. 1989. An experimental study of coevolution between the cuckoo, Cuculus canorus, and its hosts. I. Host egg discrimination. J Anim Ecol, 58:207–224.
Davies NB. 2000. Cuckoos, Cowbirds and Other Cheats. T & AD Poyser, London.
Dawkins NB, Krebs JR. 1979. Arms races between and within species. Proc R Soc B, 205:489–511.
de la Colina MA, Pompilio L, Hauber ME, Reboreda JC, Mahler B. 2012. Different recognition cues reveal the decision rules used for egg rejection by hosts of a variably mimetic avian brood parasite. Anim Cogn, 15:881–889.
Edvardsen E, Moksnes A, Røskaft E, Øien IJ, Honza M. 2001. Egg mimicry in cuckoos parasitizing four sympatric species of Acrocephalus warblers. Condor, 103:829–837.
Endler JA, 1993. The color of light in forests and its implications. Ecol Monogr, 63:1–27.
Endler JA, Milke PW Jr. 2005. Comparing entire colour patterns as birds see them. Biol J Linn Soc, 86:405–431.
Fuisz TI, de Kort SR. 2007. Habitat-dependent call divergence in the common cuckoo: is it a potential signal for assortative mating? Proc R Soc B, 274:2093–2097.
Gomez D. 2010. AVICOL, Version 4. A program to analyse spectrometric data. Available from the author upon request (dodogomez@yahoo. fr) or by download. http://sites.google.com/site/avicolprogram/.
Hargitai R, Moskát C, Bán M, Gil D, López-Rull I, Solymos E. 2010. Eggshell characteristics and yolk composition in the common cuckoo Cuculus canorus: are they adapted to brood parasitism? J Avian Biol, 41:177–185.
Hart NS, Partridge JC, Cuthill IC, Bennett ATD. 2000. Visual pigments, oil droplets, ocular media and cone photoreceptor distribution in two species of passerine bird: the blue tit (Parus caeruleus L. ) and the blackbird (Turdus merula L. ). J Comp Physiol A, 186:375–387.
Hauber ME, Sherman PW. 2001. Self-referent phenotype matching: theoretical considerations and empirical results. Trends Neurosci, 24:609–616.
Higuchi H. 1998. Host use and egg color of Japanese cuckoos. In: Rothstein SI, Robinson SK (eds) Parasitic Birds and Their Hosts. Studies in Coevolution. Oxford University Press, New York, pp 80–93.
Honza M, Moksnes A, Roskaft E, Stokke B. 2001. How are different common cuckoo Cuculus canorus egg morphs maintained? An evaluation of different hypotheses. Ardea, 89:341–352.
Honza M, Procházka P, Morongová K, Capek MC, Jelinek V. 2011. Do nest light conditions affect rejection of parasitic eggs? A test of the light environment hypothesis. Ethology, 117:539–546.
Igic B, Cassey P, Grim T, Greenwood DR, Moskát C, Rutila J, Hauber ME. 2012. A shared chemical basis of avian hostparasite egg colour mimicry. Proc R Soc B, 279:1068–1076.
Johnsgard PA. 1997. The Avian Brood Parasites: Deception at the Nest. Oxford University Press, New York.
Kelber A, Vorobyev M, Osorio D. 2003. Animal colour vision ― behavioural tests and physiological concepts. Biol Rev, 78:81–118.
Kilner RM. 2006. The evolution of egg colour and patterning in birds. Biol Rev, 81:383–406.
Krüger O. 2007. Cuckoos, cowbirds and hosts: adaptations, trade-offs and constraints. Phil Trans R Soc B, 362:1879–1886.
Langmore NE, Stevens M, Maurer G, Kilner RM. 2009. Are dark cuckoo eggs cryptic in host nests? Anim Behav, 78:461–468.
Leisler B, Schulze-Hagen K. 2011. The Reed Warblers: Diversity in a Uniform Bird Family. KNNV Uitgeverij.
Lotem A, Nakamura H, Zahavi A. 1992. Rejection of cuckoo eggs in relation to host age: a possible evolutionary equilibrium. Behav Ecol, 3:128–132.
Lotem A, Nakamura H, Zahavi A. 1995. Constrains on egg discrimination and cuckoo-host co-evolution. Anim Behav, 49:1185–1209.
Lovászi P, Moskát C. 2004. Break-down of arms race between the red-backed shrike (Lanius collurio) and common cuckoo (Cuculus canorus). Behaviour, 141:245–262.
Moksnes A, Røskaft E, Bicik V, Honza M, Øien IJ. 1993. Cuckoo Cuculus canorus parasitism on Acrocephalus warblers in Southern Moravia in the Czech Republic. J Ornithol, 134:425–434.
Moksnes A, Røskaft E, Braa AT, Korsnes L, Lampe HM, Pedersen HC. 1991. Behavioural responses of potential hosts towards artificial cuckoo eggs and dummies. Behaviour, 116:64–89.
Moksnes A, Røskaft E. 1995. Egg-morphs and host preference in the common cuckoo (Cuculus canorus): an analysis of cuckoo and host eggs from European museum collections. J Zool Lond, 236:625–648.
Møller AP, Antonov A, Stokke BG, Fossøy F, Moksnes A, Røskaft E, Takasu F. 2011. Isolation by time and habitat and coexistence of distinct host races of the common cuckoo. J Evol Biol, 24:676–684.
Molnár B. 1944. The cuckoo in the Hungarian plain. Aquila, 51:100–112.
Moskát C, Avilés JM, Bán M, Hargitai R, Zölei A. 2008a. Experimental support for the use of egg uniformity in parasite egg discrimination by cuckoo hosts. Behav Ecol Sociobiol, 62:1885–1890.
Moskát C, Hansson B, Barabás L, Bártol I, Karcza Z. 2008b. Common cuckoo Cuculus canorus parasitism, antiparasite defence and gene flow in closely located populations of great reed warblers Acrocephalus arundinaceus. J Avian Biol, 39:663–671.
Moskát C, Honza M. 2000. Effect of nest and nest site characteristics on the risk of cuckoo Cuculus canorus parasitism in the great reed warbler Acrocephalus arundinaceus. Ecography, 23:335–341.
Moskát C, Honza M. 2002. European Cuckoo Cuculus canorus parasitism and host's rejection behaviour in a heavily parasitized great reed warbler Acrocephalus arundinaceus population. Ibis, 144:614–622.
Moskát C, Székely T, Cuthill IC, Kisbenedek T. 2008c. Hosts' responses to parasitic eggs: which cues elicit hosts' egg discrimination? Ethology, 114:186–194.
Muñoz AR, Altamirano M, Takasu F, Nakamura H. 2007. Nest light environment and the potential risk of common cuckoo (Cuculus canorus) parasitism. Auk, 124:619–627.
Nakamura H, Kubota S, Suzuki R. 1998. Coevolution between the common cuckoo and its major hosts in Japan. In: Rothstein SI, Robinson SK (eds) Parasitic Birds and Their Hosts. Oxford University Press, New York, pp 94–112.
Nakamura H. 1990. Brood parasitism by the cuckoo Cuculus canorus in Japan and the start of new parasitism on the azurewinged magpie Cyanopica cyana. Japn J Ornithol, 39:1–18.
Osorio D, Vorobyev M. 1996. Colour vision as an adaptation to frugivory in primates. Proc R Soc B, 263:593–599.
Payne R. 2005. Cuckoos, Cuculidae. Oxford University Press, Oxford.
Polacikova L, Grim T. 2010. Blunt egg pole holds cues for alien egg discrimination: experimental evidence. J Avian Biol, 41:111–116.
Røskaft E, Moksnes A, Stokke BG, Bicik V, Moskát C. 2002. Agression to dummy cuckoos by potential European cuckoo hosts. Behaviour, 139:613–628.
Røskaft E, Takasu F, Moksnes A, Stokke BG. 2006. Importance of spatial habitat structure on establishment of host defenses against brood parasitism. Behav Ecol, 17:700–708.
Rothstein SI. 1990. A model system of coevolution: avian brood parasitism. Ann Rev Ecol Syst, 21:481–508.
Southern HN. 1954. Mimicry in cuckoos' eggs. In: Huxley J, Hardy AC, Ford EB (eds) Evolution as a Process. Allen & Unwin, London, pp 219–232.
Spottiswoode CN, Stevens M. 2010. Visual modelling shows that avian host parents use multiple visual cues in rejecting parasitic eggs. PNAS, 107:8672–8676.
Stoddard MC, Stevens M. 2010. Pattern mimicry of host eggs by the common cuckoo, as seen through a bird's eye. Proc R Soc B, 277:1387–1393.
Stokke BG, Moksnes A, Røskaft E. 2002. Obligate brood parasites as selective agents for evolution of egg appearance in passerine birds. Evolution, 56:199–205.
Stokke BG, MoksnesA, Røskaft E. 2005. The enigma of imperfect adaptations in hosts of avian brood parasities. Ornithol Sci, 4:17–29.
Takasu F, Moskát C, Muñoz AR, Imanishi S, Nakamura H. 2009. Adaptations in the common cuckoo (Cuculus canorus) to host eggs in a multiple-hosts system of brood parasitism. Biol J Linn Soc, 98:291–300.
Takasu F, Moskát C. 2011. Modeling the consequence of increased host tolerance toward avian brood parasitism. Popul Ecol, 53:187–193.
Takasu F. 1998. Modelling the arms race in avian brood parasitism. Evol Ecol, 12:969–987.
Takasu F. 2003. Co-evolutionary dynamic of egg apperance in avian brood parasitism. Evol Ecol Res, 5:345–362.
Trnka A, Prokop P. 2011. Polygynous great reed warblers Acrocephalus arundinaceus suffer more cuckoo Cuculus canorus parasitism than monogamous pairs. J Avian Biol, 42:192–195.
Underwood TJ, Sealy SG. 2002. Adaptive significance of egg coloration. In: Deeming DC (ed) Avian Incubation. Oxford University Press, Oxford, pp 280–298.
Vikan JR, Fossøy F, Huhta E, Moksnes A, Røskaft E, Stokke BG. 2011. Outcomes of brood parasite–host interactions mediated by egg matching: common cuckoos Cuculus canorus versus Fringilla finches. PLoS ONE, 6: e19288.
Vorobyev M, Osorio D. 1998. Receptor noise as a determinant of colour thresholds. Proc R Soc B, 265:351–358.
Yang C, Liang W, Cai Y, Shi S, Takasu F, Møller AP, Antonov A, Fossøy F, Moksnes A, Røskaft E, Stokke BG. 2010. Coevolution in action: disruptive selection on egg colour in an avian brood parasite and its host. PLoS ONE, 5: e10816.


 DownLoad:
DownLoad:





 Email Alerts
Email Alerts RSS Feeds
RSS Feeds