Sabah Mushtaq Puswal, Guo Zhong, Xuan Zhang, Longwu Wang, Wei Liang. 2024: Common Cuckoo (Cuculus canorus) nestlings adapt their begging behavior to the host signal system. Avian Research, 15(1): 100195. DOI: 10.1016/j.avrs.2024.100195
Citation:
Sabah Mushtaq Puswal, Guo Zhong, Xuan Zhang, Longwu Wang, Wei Liang. 2024: Common Cuckoo (Cuculus canorus) nestlings adapt their begging behavior to the host signal system. Avian Research, 15(1): 100195. DOI: 10.1016/j.avrs.2024.100195
Sabah Mushtaq Puswal, Guo Zhong, Xuan Zhang, Longwu Wang, Wei Liang. 2024: Common Cuckoo (Cuculus canorus) nestlings adapt their begging behavior to the host signal system. Avian Research, 15(1): 100195. DOI: 10.1016/j.avrs.2024.100195
Citation:
Sabah Mushtaq Puswal, Guo Zhong, Xuan Zhang, Longwu Wang, Wei Liang. 2024: Common Cuckoo (Cuculus canorus) nestlings adapt their begging behavior to the host signal system. Avian Research, 15(1): 100195. DOI: 10.1016/j.avrs.2024.100195
Ministry of Education Key Laboratory for Ecology of Tropical Islands, Key Laboratory of Tropical Animal and Plant Ecology of Hainan Province, College of Life Sciences, Hainan Normal University, Haikou, 571158, China
b.
School of Life Sciences, Guizhou Normal University, Guiyang, 550001, China
Funds: This work was supported by the National Natural Science Foundation of China (Nos. 32270526 to W.L. and 32260253 to L.W.). W.L. was supported by the specific research fund of The Innovation Platform for Academicians of Hainan Province. S.M.P. was supported by the Hainan Province Postdoctoral Research Project
Common Cuckoos (Cuculus canorus) dependent on parental care for post-hatching demonstrate an intriguing ability to modify their begging vocalizations to ensure maximum care and resources from their interspecific foster parents. Here, we compared begging calls of the Common Cuckoo nestlings fed by four host species, the Grey Bushchat (Saxicola ferreus), Siberian Stonechat (Saxicola maurus), Daurian Redstart (Phoenicurus auroreus), and Oriental Magpie-robin (Copsychus saularis). Results showed that begging calls of the stonechat–, redstart–, and robin–cuckoo resemble those of host species' nestlings in various aspects like low frequency, high frequency, frequency bandwidth and peak frequency, while the bushchat–cuckoo chicks' begging calls were only comparable to their host species in terms of how long they lasted and their peak frequency. In addition, cuckoo nestlings raised in different host nests displayed significant variations in their begging calls in low and peak frequency. This study suggests that cuckoo nestlings do not mimic host species nestlings' begging calls throughout the nestling period, but may tune their begging calls according to host species, while begging calls vary with cuckoo and host species nestlings' ages. Future research should study the parents' reactions to these calls in different host species for a better understanding of the mechanisms underlying such adaptations.
Parasites manipulating a host to raise their offspring engage in brood parasitism (Feeney et al., 2012). Brood parasitism means the hosts have additional parental care responsibility following parasitic females laying their eggs in the host species' nest (Ruiz-Raya et al., 2017). Parasitism, when successful, has negative consequences on the host as it causes a decline in the host's fitness due to the loss of some or all host progeny (Kilner et al., 2004). This reproductive cost imposed on the host by parasitic species leads to defensive host adaptations, resulting in a co-evolutionary arms race between parasites and hosts that results in host resistance to parasitism. In contrast, the parasites adopt coping mechanisms as a response to the defenses put up by the hosts (Dawkins and Krebs, 1979; Davies, 2000; Wang et al., 2020). One of the best studied example of this evolutionary behavior is the rejection of foreign eggs by host species and the mimicry of hosts' eggs by parasitic females (Yang et al., 2014, 2020).
Likewise, some host species may accept the eggs but reject the parasite chicks (Sato et al., 2010; Tokue and Ueda, 2010), leading to some cuckoo species imitating host chicks to increase the chance for parental acceptance and care by the host (Langmore et al., 2008; Jamie et al., 2020). Theoretically, this evolutionary behavior of differentiating chicks and rejecting cuckoo chicks based on imprinting carries a significant risk since the host could erroneously imprint on a parasitic chick in their first brood, causing them always to reject their chicks after that (Lotem, 1993). However, some host species have evolved mechanisms that allow them to reject parasitic cuckoo chicks without the need for recognition, demonstrating the magnitude of behavioral adaptation by the host species (Grim, 2007; Langmore et al., 2008).
In avian species, nestlings usually depend on the adults, hence they use begging displays, a combination of visual and acoustic signals to communicate their needs to their care-giving adults; in turn, the care-giving adults use these signals to determine how much food they should provide to each offspring (Leonard and Horn, 2001; Glassey and Forbes, 2002; Leonard et al., 2003; Rojas Ripari et al., 2021). Because these signals (begging calls) vary among species and play a major role in signaling offspring identity (Beecher et al., 1981), they may act as cue for nestling rejection. This coevolutionary behavior is best explained through the Superb Fairy-wrens (Malurus cyaneus), which have a low probability of rejecting the Horsefield Bronze Cuckoo (Chalcites basalis) chicks whose calls resemble to their own offsprings but have a high probability of rejecting Shining Bronze Cuckoo (Chalcites lucidus) chicks whose calls do not (Langmore et al., 2003). Parasitic chicks are thus expected to generate begging signals that "tune into" the parent-offspring communication system (Gloag and Kacelnik, 2013; Rojas Ripari et al., 2021). For example, Great Spotted Cuckoo (Clamator glandarius) nestlings make acoustic adaptations by adjusting the number of notes in their calls depending on their feeding needs with the rearing host (Roldán et al., 2013). Similarly, Greyish Baywing (Agelaioides badius) parents use acoustic signals to differentiate their fledglings from foreign ones; in turn, brood parasitic Cowbird (Molothrus rufoaxillaris) nestlings prevent host discrimination by mimicking begging calls (Fraga, 1998; De Mársico et al., 2012; Lama et al., 2022).
Acoustic tuning into the host environment has also been shown to occur in the Common Cuckoo (Cuculus canorus), an evictor parasite that targets a range of host species each with different traits (Honza et al., 2007; Yang et al., 2012, 2020). Studies have shown that Common Cuckoo nestlings had a more rapid calling rate in Dunnock (Prunella modularis) host nests than in Reed Warbler (Acrocephalus scirpaceus) nests to fit in the host system despite growing at the same rate (Butchart et al., 2003). Madden and Davies (2006) found that begging calls by the Common Cuckoo nestlings raised naturally by Reed Warblers and Dunnocks have variations in their acoustic features. Other variations are best explained using the cross-fostered Reed Warbler cuckoos, which have been reported to modify their syllable duration, minimum frequency, and bandwidth to mimic those of Dunnock cuckoo nestlings. This suggests that cuckoos have better acoustic adaptation to the host environment, which reduces the probability of rejection while increasing their chance of getting fed by the host due to their high degree of host specialization and great ability to attuning into the host system (Jamie and Kilner, 2017).
Nevertheless, the results of earlier studies on cuckoo nestlings' calls are paradoxical where some suggested that their begging calls are plastic whereas others argue that cuckoo nestling calls are similar in different hosts' nests and they use general begging call features to obtain food from foster parents. For example, Soler (2017) found that call structure differed between cuckoos in four host species' nests, but not call rates. On the contrary, Samaš et al. (2020) found that Common Cuckoo nestlings do not mimic begging calls of two closely related warbler species, but changes in begging calls such as a narrowed frequency band, shortened call bandwidth, reduced syllable duration, and increased call rate were better explained through nestling ages. Noh et al. (2021) also found that the differences in the begging calls of Little Bronze Cuckoos (Chalcites minutillus), and their host's Large-billed Gerygones (Gerygone magnirostris) nestlings were more distinct as they got older. This suggests that parasitic nestlings do not use vocal mimicry unless there is a high probability of rejection by the host (Noh et al., 2021) and age is an important factor to consider when comparing nestling vocalizations. Furthermore, Briskie et al. (1999) found that birds subjected to high predation risk had calls with high frequency and low amplitude compared to the species with low predation risk. Therefore, call similarity by parasite nestlings may be due to other factors such as predation risk, ambient noise, and relatedness (Haskell, 1999; Boncoraglio and Saino, 2008; Leonard and Horn, 2008). In fact, several studies confirm that the host parents do not reject parasite nestlings in all cases where call types match (Redondo and Reyna, 1988; Anderson et al., 2009). Thus, begging call mimicry may be adopted to increase the level of parental care by the host and not to reduce rejection probability (Jamie and Kilner, 2017). Therefore, further research on the differences and similarities of cuckoo nestling begging calls will help create a better understanding of the evolutionary behaviors observed in brood parasitism.
In this study, we investigated begging calls of Common Cuckoo nestlings fed by four different host species, namely the Grey Bushchat (Saxicola ferreus), Siberian Stonechat (S. maurus), Daurian Redstart (Phoenicurus auroreus) and Oriental Magpie-robin (Copsychus saularis). These species have different body sizes and have partial overlap in breeding periods. We aimed to investigate whether: (1) cuckoo nestling calls differ in different host species nests; (2) cuckoo nestlings mimic the begging calls of host species nestlings throughout the nestling period. Previous studies have found that cuckoo chicks tune into different offspring-parent communication systems in nests of different host species (Madden and Davies, 2006; Jamie and Kilner, 2017). We predicted that the begging calls of cuckoo reared by different host species would differ in at least some acoustic parameters. Based on the earlier studies which have reported a significant difference in cuckoo and host species' nestlings with age (Noh et al., 2021), we also expected a difference in cuckoo and host species' call features with days. Furthermore, earlier studies have reported changes in call structure with increasing age (Butchart et al., 2003). For example, an increase in call duration did result in enhanced feeding rates (Gloag and Kacelnik, 2013), thus we also expected a similar change in call structure.
2.
Materials and methods
2.1
Study area and studied species
This research was conducted in Liuzhi District, Guizhou Province, southwest China (26°10′–14′ N, 105°13′–24′ E) from May to August 2022, during the breeding season of the Common Cuckoo and its host species. In total, the study area covers about 1800 ha, and the average elevation is 1442 m a.s.l. The area is mainly covered with forests, shrubs and farmlands. Weather conditions in the study area are subtropical monsoon, with an average annual temperature of 13.9 ℃ (summer 21 ℃) and total precipitation of 1515.55 mm (Liu et al., 2023).
In our study area the Common Cuckoo uses the Grey Bushchat, Siberian Stonechat, Daurian Redstart (Phoenicurus auroreus), Oriental Magpie-robin, Yellow-throated Bunting (Emberiza elegans), South Rock Bunting (E. yunnanensis), Crested Bunting (E. lathami) and the White Wagtail (Motacilla alba) as host species (Wan et al., 2023; Zhong et al., 2023). Among these hosts the Grey Bushchat is the primary host and the parasitism rate is 8.2% (Zhong et al., 2023).
The Grey Bushchat (14–16 g) and Siberian Stonechat (13–17 g) are insectivorous passerine birds belonging to the order Passeriformes. The former is distributed in east, south, central, northwest, and southwest China, whereas the latter breeds in southwest China (Zheng, 2023). The nests of both species are semi-open, cup-shaped, made from fine roots and grasses, lined with feathers and hairs, and usually placed on the ground. Their clutch size ranges from 4 to 6 eggs, and the nestlings usually fledge after 11–12 days (Zhong et al., 2023).
The Daurian Redstart weighs 11–20 g and the Oriental Magpie-robin weighs 29–51.3 g, respectively, belonging to the order Passeriforme. The former species is found throughout China except for Xinjiang, Tibet, and Qinghai whereas the latter species is located mostly in southern China (Zheng, 2023). Both species feed mainly on insects, but the former sometimes also forages on grasses and seeds. These two species usually nest in holes, house wall joints, bamboo clumps, and on old buildings roofs (Collar, 2020). They lay between 3 and 5 eggs, and their nestlings usually fledge after 12 days. All species breed from April to mid-August, and are commonly parasitized by the Common Cuckoo (Yang et al., 2012).
2.2
Begging call recordings
To collect call data, we systematically searched for potential areas for cuckoo hosts during the breeding season by observing host species' activities. After locating the host species' nests, we checked them daily to track hatching dates. In order to examine evidence of mimicry, we recorded calls from four hosts' nestlings at six and twelve days, and cuckoo nestlings' calls from each host species' nest at six and twelve days. There were three to five nestlings for each host species, whereas, for cuckoo nestlings, each nest had one cuckoo chick (Appendix Table S1). All cuckoo nestlings were hatched from the cuckoo–host race with blue eggs found in parasitized Grey Bushchat hosts (Zhong et al., 2023). The eggs were placed in an incubator until they hatched, and then they were moved to the nests of the host parents on the same day of hatching. The recordings were then analyzed to compare the vocalizations of the host and cuckoo nestlings. The begging calls of the host and cuckoo nestlings were recorded by using an audio recorder (Sony PCM-A10, Tokyo, Japan). The recorder was placed approximately 12–14 cm above the nest, and each recording lasted for more than 45 min (Butchart et al., 2003). All the recordings were made in WAV form at sampling rate of 44.1 kHz and 16 bits accuracy and the data were saved for further analysis. Furthermore, we installed video cameras 2 m away from the nest to record chick behavior and ensure that recordings did not interfere with the parents' feeding rate.
2.3
Acoustic analysis
For the acoustic analysis, we selected five good quality begging calls from each recording when parents visited the nests for feeding. For each call (note) we measured five acoustic variables: maximum frequency (kHz), minimum frequency (kHz), peak frequency (kHz), call duration (s) and frequency bandwidth (kHz) (Butchart et al., 2003; Noh et al., 2021). All measurements of begging calls were performed in Raven Pro 1.5 sound analysis software (Bioacoustics Research Program, 2014) by using a Hann window FFT = 512, overlap 50% and frame size 100%.
2.4
Statistical analysis
The begging calls of host species' nestlings and Common Cuckoo nestlings reared in different host species' nests were compared using linear mixed-effects models. In the first step, we compared the begging calls of four host species to examine if they differed significantly between nestling age and species. In our models, response variables were different call parameters. The models included species, nestling age, and the interaction between nestling age and species as fixed effects, and nest ID as a random effect. We used lmer function in the lmerTest package to fit the linear mixed models (Bates and Maechler, 2009). In the next step, we performed a stepwise discriminant functional analysis to determine if the model could distinguish the calls of Common Cuckoo nestlings and host species' nestlings at different stages of the nestling period (Langmore et al., 2008). The discriminant functional analysis was performed in JPM software. To examine how age affects different begging call features we performed a separate regression analysis for each host and cuckoo host race (Butchart et al., 2003). We then conducted a combined regression analysis for each cuckoo host race and host species nestlings to test how different call features change with age (Samaš et al., 2020).
To compare the begging calls of cuckoo nestlings raised in four host species' nests, we ran a linear mixed effect model with age, cuckoo species and their interaction as fixed effects and nest ID as a random factor. In the cases of significant differences between cuckoo nestling calls in different hosts' nests, they were further compared by the Tukey HSD test using the multcomp package in R. The statistical analyses were performed using R v.4.2.2 (R Development Core Team R, 2018) and the results were considered statistically significant at P <0.05.
3.
Results
Our results indicated that the host nestlings' begging calls differed significantly in duration, high frequency, frequency bandwidth and peak frequency between age (F1,38 = 6.31, P = 0.01; F1,24.1 = 6.31, P = 0.0001; F1,20.6 = 34.4, P = 0.0001; F1,117 = 9.33, P = 0.002), nestlings (F3,45.7 = 29.71, P = 0.0001; F3,29.1 = 8.31, P = 0.0003; F3,24.9 = 10.94, P = 0.0001; F3,117 = 5.35, P = 0.001), as well as by their interactions (F3,47.9 = 35.33, P = 0.0001; F3,31.2 = 6.25, P = 0.001; F3,25.6 = 8.56, P = 0.0004; F3,117 = 8.69, P = 0.0002), respectively. Furthermore, there were also differences in low frequency between species (F3,45.5 = 3.68, P = 0.01) as well as interactions between age and species (F3,46.5 = 8.69, P = 0.0001), but they were similar between days (F1,40.9 = 1.7, P = 0.19).
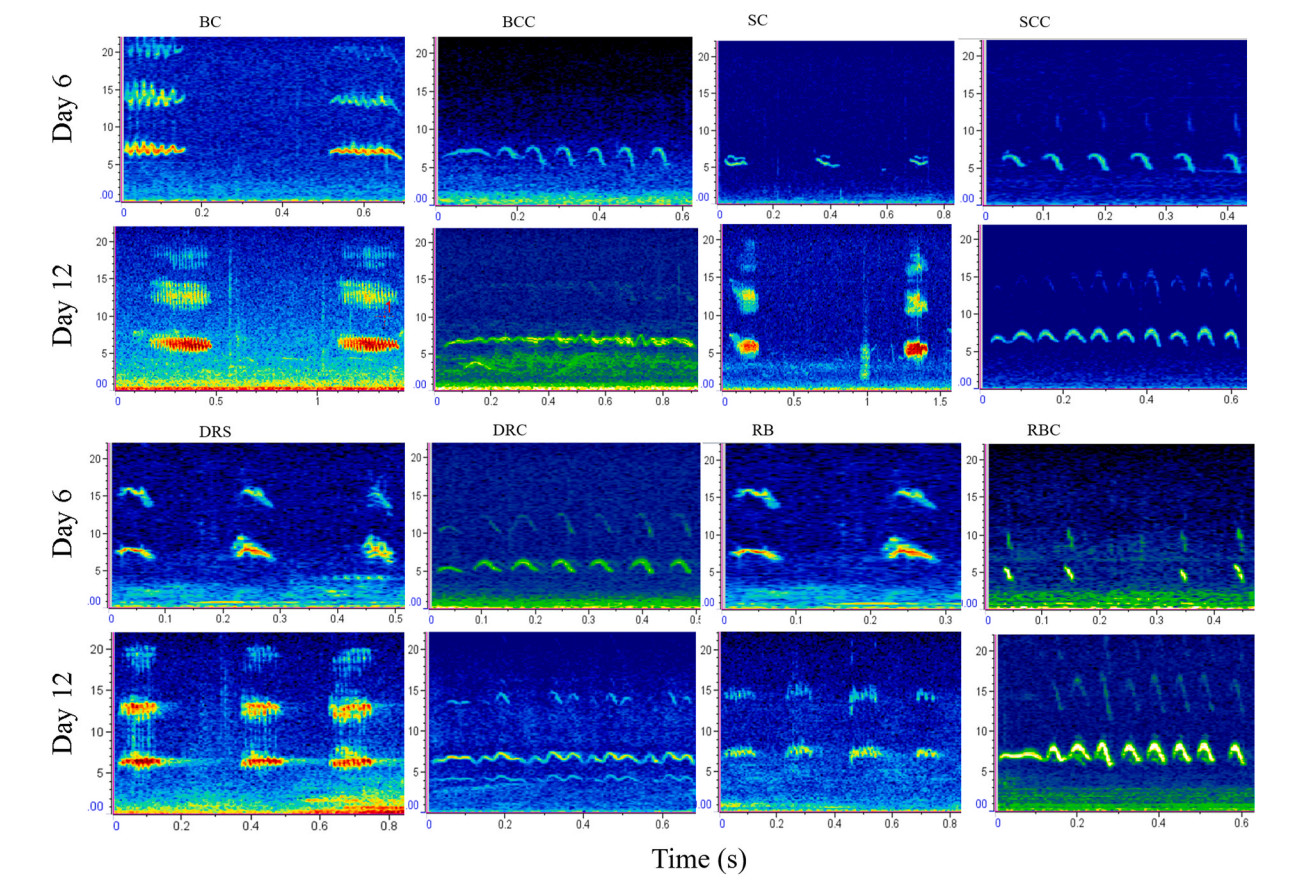
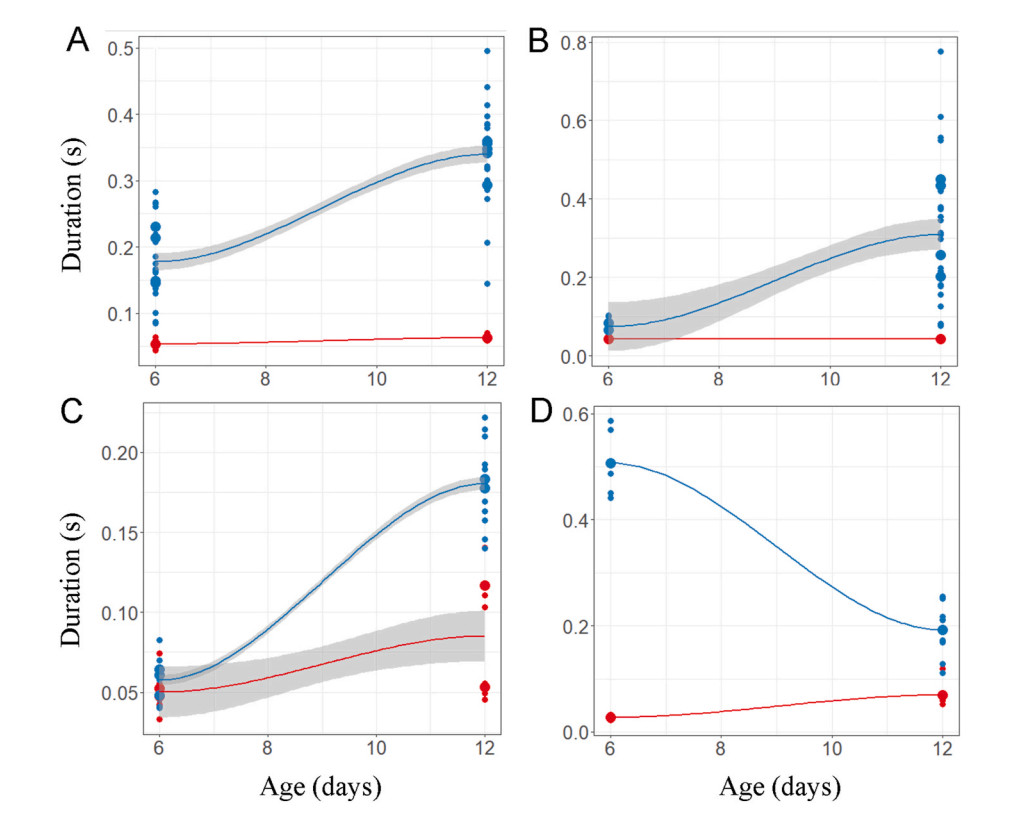
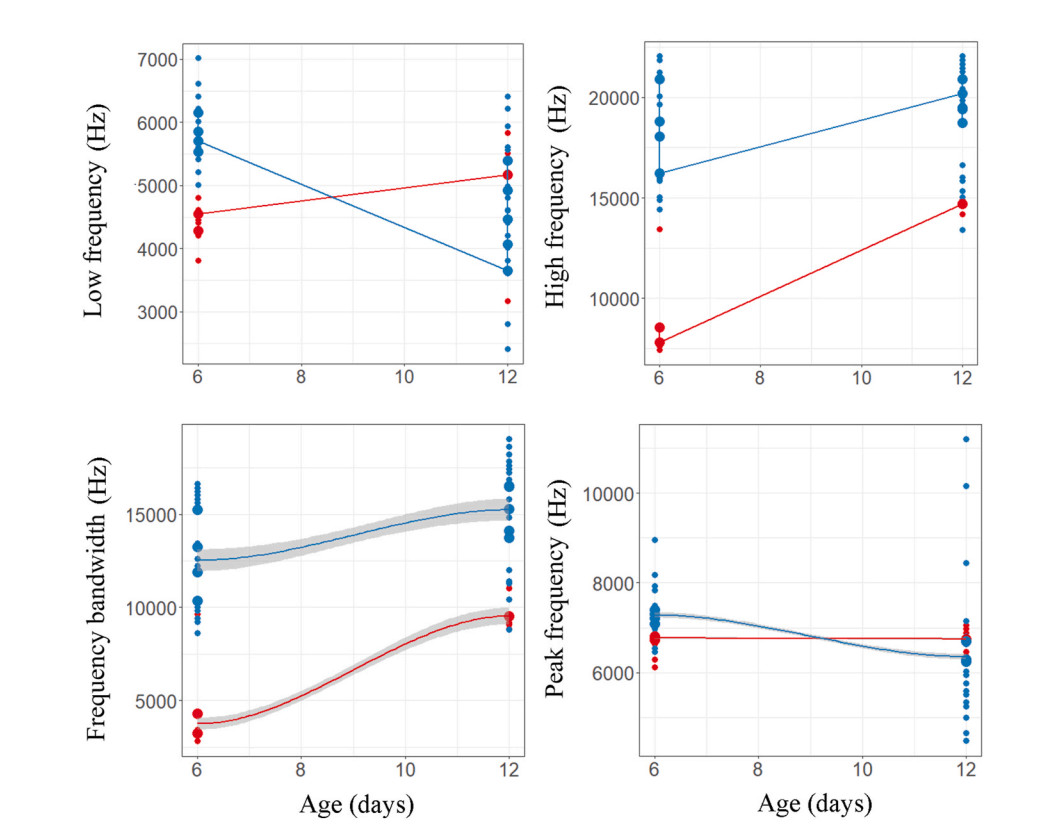
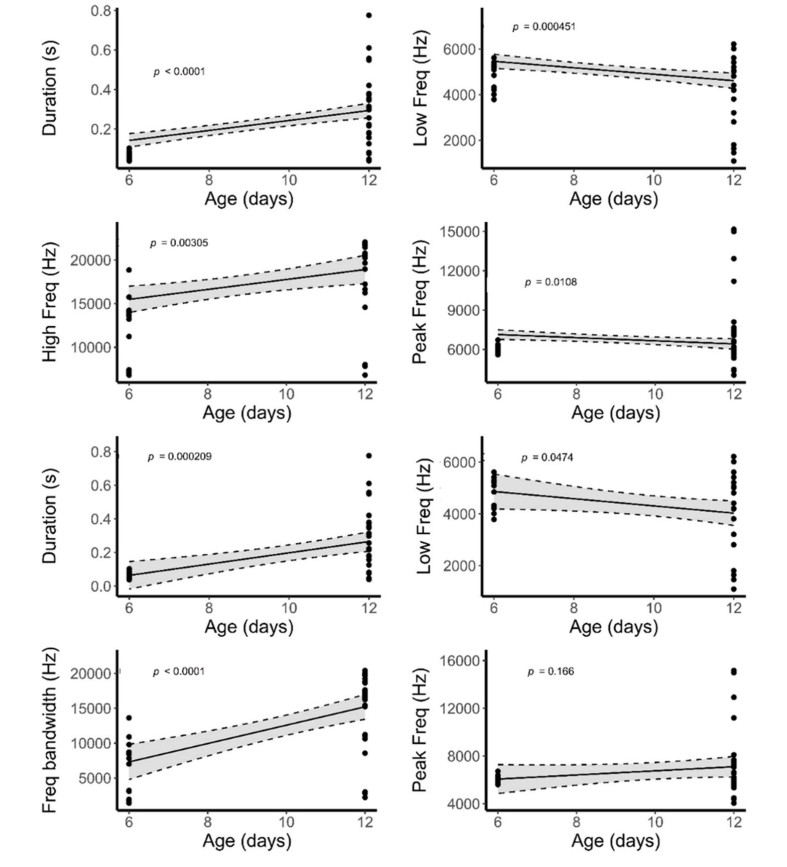
Our linear mixed effect model indicated that the begging calls of Common Cuckoo nestlings are significantly different from those of Grey Bushchat nestlings on all days and the interaction between nestling age and species were also different, indicating that changes in call structure with age differ between bushchat cuckoo and bushchat nestlings (Fig. 3 and Table 1). In contrast, duration and peak frequency of begging calls of Common Cuckoo nestlings were similar to those of bushchat nestlings, the interaction between nestling age and species was also not significant for peak frequency, but for begging duration age and interaction were different (Fig. 3 and Table 1). The call features of bushchat cuckoos and host species changed significantly as time progressed (Table 2), and when the data from both species were combined in a single regression analysis, the low frequency and peak frequency of calls decreased significantly over time, whereas the other three call variables increased significantly with age (Fig. 7). Consistent with this, a discriminant functional analysis was successful in identifying the calls of bushchat cuckoo and bushchat nestlings with high accuracy when nestlings were 6 days old (all call parameters: Wilk's = 0.03, Exact F5,9 = 44.4, P = 0.001). By contrast, it failed to discriminate the bushchat cuckoo calls based on the low and peak frequency of calls (step-wise procedure: Wilk's = 0.87, Exact F1,8 = 1.13, P = 0.31; Wilk's = 0.84, Exact F1,8 = 1.41, P = 0.26), respectively, when nestlings were 12 days old (Fig. 3).
Table
1.
Analysis of variance of age matched begging call features of cuckoo nestlings and host species' nestlings raised in four host species' nests (SE refers to standard error and significant results are shown in bold).
Table
2.
Linear regression analysis of begging calls of cuckoo nestlings and four host species nestlings against age (SE refers to standard error and significant results are shown in bold).
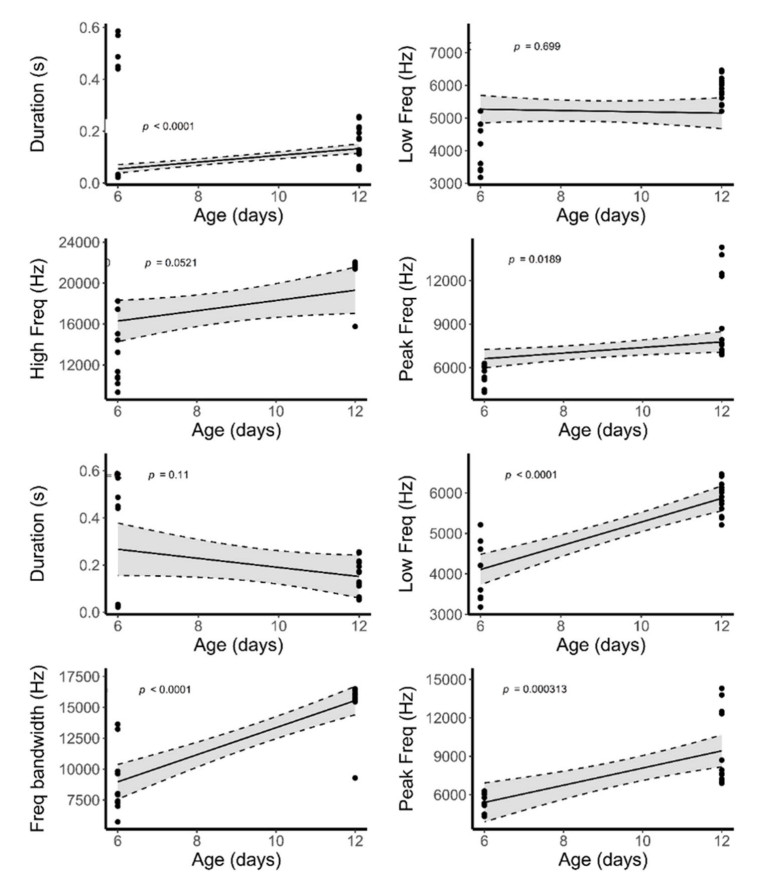
According to the linear mixed model, the begging calls of the stonechat cuckoo closely resemble those of the host species in duration, low frequency and peak frequency (Table 1), whereas high frequency and frequency bandwidth varied significantly with age and the interaction between nestlings and age was also different for frequency bandwidth (Table 1). The duration, high frequency, and frequency bandwidth of stonechat cuckoos and stonechat nestlings increased with age (Table 2), whereas low frequency decreased, and peak frequency did not change (Fig. 7). A discriminant analysis failed to differentiate the calls of parasitic cuckoo and stonechat nestlings at 6 days of age on the basis of high frequency (Wilk's = 0.93, Exact F1,13 = 0.95, P = 0.35), frequency bandwidth (Wilk's = 0.98, Exact F1,13 = 0.21, P = 0.64, and peak frequency (Wilk's = 0.93, Exact F1,13 = 0.88, P = 0.36) (Fig. 4). However, it has successfully assigned calls to both species on the basis of duration (Wilk's = 0.71, Exact F1,28 = 10.94, P = 0.002), low frequency (Wilk's = 0.77, Exact F1,28 = 8.09, P = 0.008), high frequency (Wilk's = 0.61, Exact F1,28 = 17.8, P = 0.0002) and frequency bandwidth (Wilk's = 0.54, Exact F1,28 = 23.4, P = 0.001) whereas peak frequency of calls remained non-significant (Wilk's = 0.99, Exact F1,28 = 0.19, P = 0.66) when nestlings were 12 days old (Fig. 4).
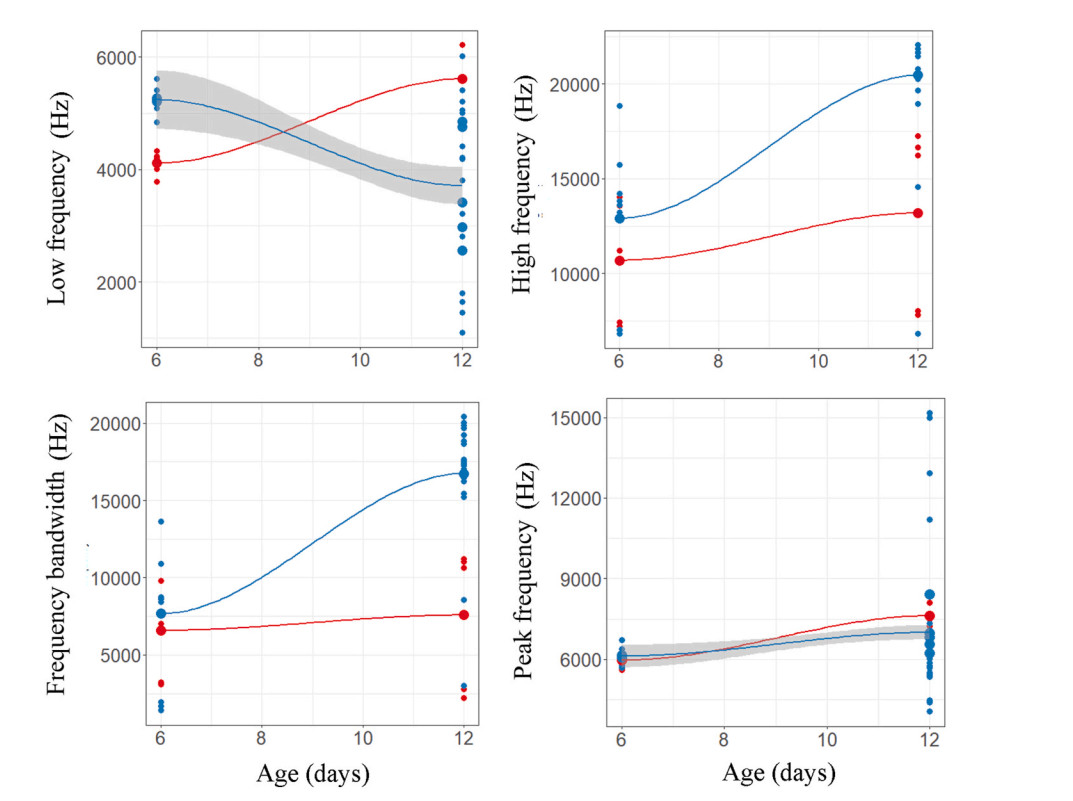
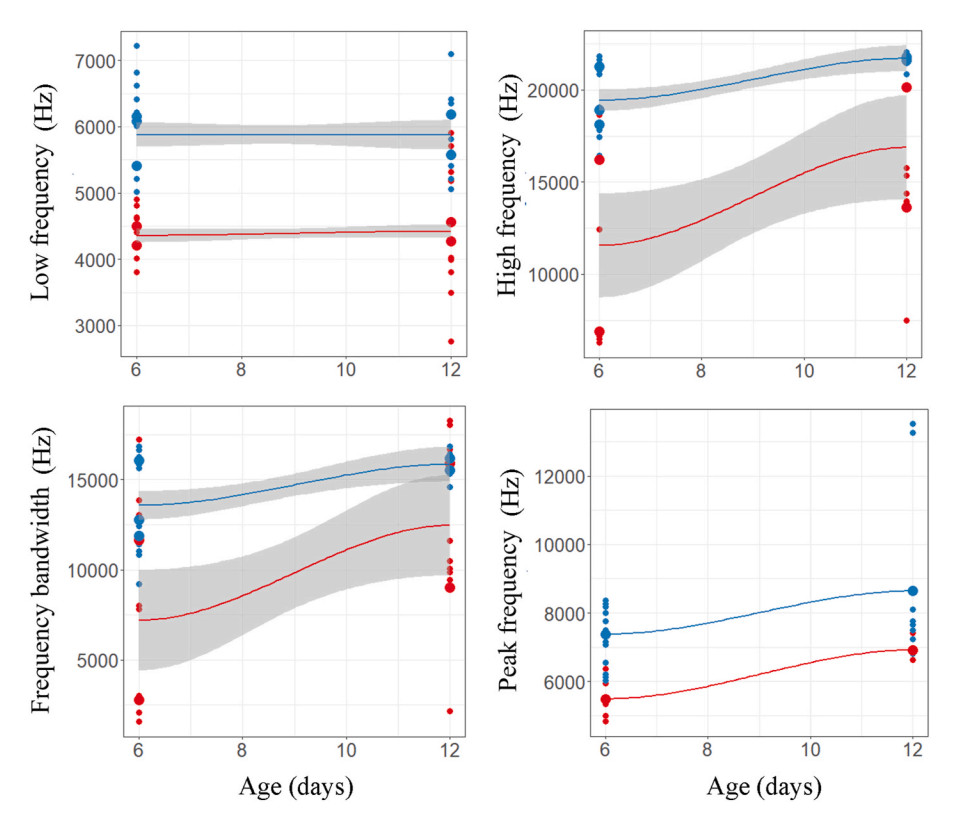
We found significant differences in peak frequency of calls between nestlings of redstart cuckoos and host species, and the interaction between nestling age and species was non-significant (Table 1). Duration of calls remained same between nestlings; however, it varied significantly with age and the interaction between nestlings and age was also significant (Table 1). The other variables, however, did not vary between species or days (Table 1). When nestlings are 6 days old, redstart cuckoo's begging calls appear similar to host species' calls in some of the acoustic parameters; however, as they grow older, their call structures became different from host species (Fig. 1 and Table 2). A discriminant analysis could not differentiate their calls by call duration (Wilk's = 0.96, Exact F1,18 = 0.72, P = 0.40) or frequency bandwidth (Wilk's = 0.94, Exact F1,18 = 1.04, P = 0.32) when redstart cuckoo nestlings were young (Figs. 2C and 5). By contrast, it could best distinguish between them on 12 days of age by using call duration (Wilk's = 0.32, Exact F1,18 = 37.2, P = 0.0001), low frequency (Wilk's = 0.55, Exact F1,18 = 14.32, P = 0.001), high frequency (Wilk's = 0.65, Exact F1,18 = 9.46, P = 0.006) and peak frequency (Wilk's = 0.79, Exact F1,18 = 4.51, P = 0.04) (Figs. 2C and 5).
Figure
1.
Spectrograms of the begging calls of cuckoo nestlings: bushchat cuckoo (BCC), stonechat cuckoo (SCC), daurian redstart cuckoo (DRC) and robin cuckoo (RBC) raised in bushchat (BC), stonechat (SC), daurian redstart (DRS) and robin (RB) nests on six and 12 days. X-axis represents time in seconds whereas Y-axis represents frequency in kilohertz (kHz).
Figure
2.
Duration of begging calls of cuckoo nestlings (red) and host species' (A) bushchat (B) stonechat (C) daurian redstart (D) robin nestlings (blue) in relation to their age.
Figure
5.
Duration and frequency parameters of begging calls of Daurian Redstart cuckoo (red) and Daurian Redstart (blue) nestlings in relation to their age.
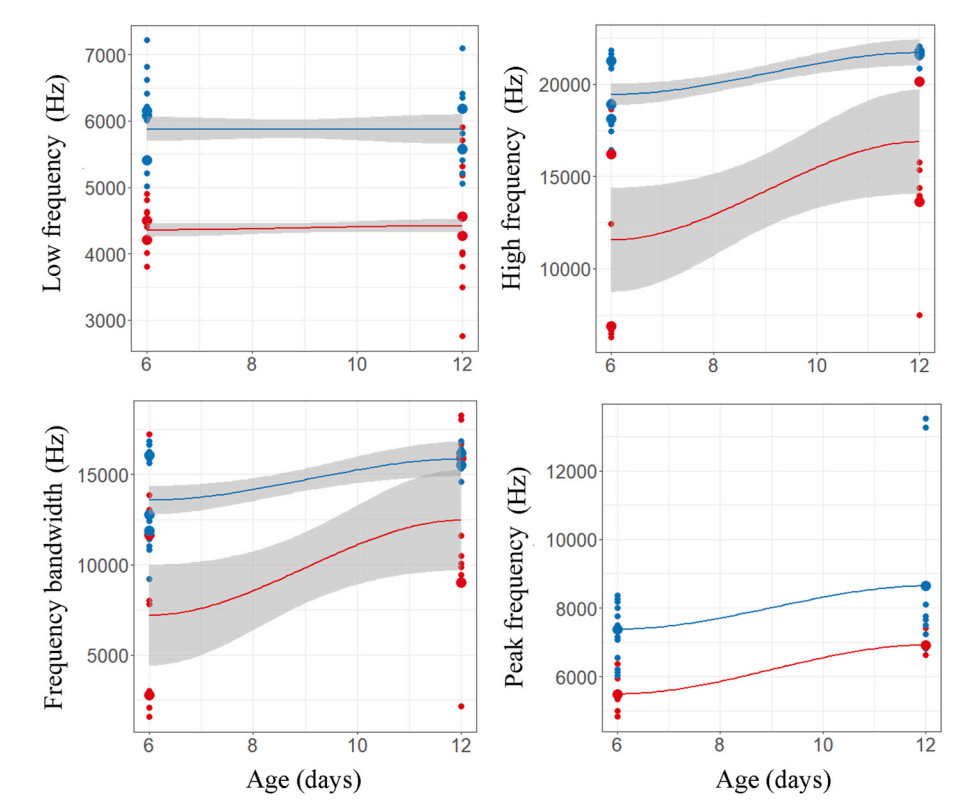
There were no differences between the begging calls of the robin cuckoo and the nestling calls of the host species in low frequency, high frequency, or bandwidth (Table 1); in contrast, the peak frequency of calls differed significantly with age (Table 1). Likewise, duration of the calls varied significantly between nestlings of both species and the interaction between them was also significant. We found that begging calls of both species changed as they aged (Figs. 1 and 8 and Table 2). A discriminant analysis could distinguish the calls of robin cuckoo and robin nestlings with more than 90% accuracy in the first half of the nestling period (all call parameters: Wilk's = 0.01, Exact F5,4 = 56.4, P = 0.0008) (Fig. 6). However, this accuracy dropped, and discriminant analysis failed to discriminate against begging calls in the latter half of the nestling growth period on the basis of low frequency (Wilk's = 0.98, Exact F1,13 = 0.24, P = 0.62), high frequency (Wilk's = 0.92, Exact F1,13 = 1.07, P = 0.31), and frequency bandwidth (Wilk's = 0.95, Exact F1,13 = 0.65, P = 0.43) (Fig. 6).
Figure
6.
Duration and frequency parameters of begging calls of robin cuckoo (red) and robin (blue) nestlings in relation to their age.
Figure
7.
Regression plots of begging call features of bushchat and bushchat cuckoo nestlings (upper four plots) and stonechat and stonechat cuckoo nestlings (lower four plots) against age.
Figure
8.
Regression plots of begging call features of Daurian Redstart and Daurian Redstart cuckoo nestlings (upper four plots) and robin and robin cuckoo nestlings (lower four plots) against age.
Based on the linear mixed effect model, the begging calls of cuckoos reared in different host species nests vary significantly from each other in low (age: F1,47 = 36.3, P = 0.0001; cuckoo nestlings: F3,47 = 7.52, P = 0.0003; age × cuckoo nestlings F3 = 9.11, P = 0.0001; Table S2) and peak frequency (age: F1,4.9 = 85.8, P = 0.0002; cuckoo nestlings: F3,14.7 = 25.03, P = 0.0001; age × cuckoo nestlings: F3,5.7 = 14.4, P = 0.004) of calls (Table S3). The other parameters high frequency (days: F1,3.1 = 17.3, P = 0.02; cuckoo nestlings: F3,5.6 = 1.61, P = 0.28; age × cuckoo nestlings: F32.51, P = 0.22) and frequency bandwidth (age: F1,3.2 = 11.9, P = 0.03; cuckoo nestlings: F3,6.1 = 0.6, P = 0.62; age × cuckoo nestlings: F31.34, P = 0.39) differ among days but did not differ among nestlings. The duration of the calls, however, remained the same (age: F1,2.8 = 1.8, P = 0.29; nestlings: F3,2.6 = 4.28, P = 0.15; age × cuckoo nestlings: F3,2.1 = 5.4, P = 0.14).
4.
Discussion
Our results indicated that vocalizations of stonechat–, redstart–, and robin–cuckoo exhibit similarities in some frequency parameters with their respective host species' nestlings, while the bushchat cuckoo mirrors its host primarily in duration and peak frequency. Further, temporal shifts in the vocal characteristics were evident across the four cuckoo variants and within their respective host species.
To ensure they get enough care and food from their interspecific foster parents, nestlings of altricial birds adjust their begging calls to match the host's parent-offspring communication style (Davies, 2011). While some species have evolved to produce louder and faster calls than their hosts' young (Butchart et al., 2003), others have adopted a strategy of closely imitating the intricate vocal structure of their host nestlings (Rojas Ripari et al., 2019). Our research showed that some features of cuckoo nestlings' begging calls were related to the host species that reared them. Perhaps various hosts respond to different call features, resulting in cuckoo nestlings emphasizing these traits instead of merely copying the complete vocal profile of their hosts' young (Madden and Davies, 2006). Earlier investigations have shown that vocal mimicry of hosts is essential for survival in the nests of hosts exhibiting chick rejection behavior (Butchart et al., 2003; Noh et al., 2021); however, based on experiments in the field and our personal observations, the studied species in this study do not reject but feed parasitic nestlings well. In such scenarios, the primary driver behind vocal plasticity might not be purely survival-based, but instead, it may be a mechanism to integrate into the parent–offspring communication dynamics seamlessly (Jamie and Kilner, 2017). For instance, Ursino et al. (2018) discovered that playing back the imitative calls of Screaming Cowbirds (Molothrus rufoaxillaris) led to increased feeding in the same way as playing back the begging calls of the host's offspring (Ursino et al., 2018). Similarly, even when there is no threat of being expelled by their hosts, both the Pin-tailed (Vidua macroura) and Broad-tailed Paradise Whydahs (V. obtusa) young still imitate the calls of their host's nestlings. This suggests that imitating begging calls is crucial for ensuring adequate care rather than to avoid being evicted from the nest (Schuetz, 2005; Jamie et al., 2020). In fact, if a foreign chick does not adapt its calls to match the feeding habits of the host, it may starve to death (Kilner et al., 1999).
Furthermore, our stepwise discriminant analysis indicated that by the 12th day of their life cycle, the calls of the bushchat cuckoo closely resembled their hosts in low and peak frequency. Likewise, the begging calls of robin and robin cuckoo were similar in low, high and frequency bandwidth. In contrast, rewind to the sixth day, these similarities become much less obvious. Such observations lead to intriguing speculations suggesting that different host species have evolved distinct preferences for specific begging vocalizations based on their evolutionary history and ecological pressures. Thus, such preferences could act as triggers, prompting parasitic nestlings to modulate their calls to maximize the allocation of resources from their foster parents (McLean and Waas, 1987; Gloag and Kacelnik, 2013). It is consistent with previous studies that highlight the remarkable plasticity of nestlings, adapting their begging calls in response to environmental cues and parental feedback (West and King, 1988; Kedar et al., 2000). Alternatively, some bird species have similar calls due to their environment or threats they face (Haskell, 1999; Leonard and Horn, 2008); however, this is not necessarily true for the species we studied. Because two of them nest on the ground, and the others in holes or walls, this means that at least two face similar threats from predators. Therefore, the most convincing reason for their call similarities is likely to match the host bird's preferred calls.
Our linear mixed model revealed that low and peak frequency of calls differ across cuckoo–host races. These findings are consistent with previous research, emphasizing the profound influence of the nurturing environment of host species on the vocal structures exhibited by cuckoo nestlings (Madden and Davies, 2006; Soler, 2017). Tuero et al. (2016) discovered that the begging patterns of cowbird chicks varied based on the host species raising them, even though they had similar condition and hunger levels (Tuero et al., 2016). Kölliker et al. (2000) further supported this environmental influence on begging behavior. By switching Great Tit (Parus major) chicks among different nests, they determined that their shared upbringing environment influenced the energy intensity variations in nestling calls. This suggests that nestlings adjust their call patterns in response to how their parents feed them (Kölliker et al., 2000). Jamie and Kilner (2017) indicated that female cuckoos exhibit genetic ploymorphic norm, suggesting they have ability to adjust their begging calls based on the environment of their hosts, so that even when brought up by the same host species, different cuckoo races may still slightly vary their calls. It is important to note that while earlier studies on cuckoo nestling calls, like Davies et al. (2006), used cross-fostering techniques, our research did not. Therefore, our findings could be influenced by the limited number of samples, as pointed out by Samaš et al. (2020). Our results indicated that as cuckoo nestlings and their host species age, there are changes in their call structures. This aligns with previous research indicating that as nestlings age and become hungrier, there is a noticeable variation in their calls' length, bandwidth, and maximum frequency (Butchart et al., 2003; Magrath et al., 2010; Samaš et al., 2020). Some playback experiments that utilized artificial calls did not find a link between call frequency parameters and feeding efforts; however, an increase in the number of syllables or prolonged call durations did result in enhanced feeding rates (Gloag and Kacelnik, 2013). This underlines the idea that among Passerines, the length of the call acts as an indicator of the nestling's hunger, leading to increased feeding (Leonard and Horn, 2001). While past studies highlighted that heavier species tend to produce calls at reduced frequencies (García et al., 2014; Mikula et al., 2021), our findings showed an increase in high-frequency calls as all species aged. This might be attributed to the louder calls made by older nestlings, which aligns with previous research showing a positive relationship between fundamental frequency and vocal effort in humans (Liénard and Di Benedetto, 1999).
In conclusion, the call patterns of cuckoo nestlings resemble those of their feeding host species in some of the parameters, indicating adaptations based on the host's environment. Even though there are discrepancies across various studies, our findings showed significant difference in low and peak frequency of calls when analyzing all cuckoo nestlings collectively. Future research should utilize larger sample sizes and consider cross-fostering cuckoo nestlings for more host species to compare their begging calls comprehensively. Moreover, to assess if nestlings adjust their begging calls to receive more food, it is imperative to study the response behavior of foster parents to these calls. Perhaps this would offer a better understanding of cuckoo nestlings' strategies to influence the care they receive from their foster parents.
Ethics statement
The experiments complied with current laws in China. The experimental procedures were performed per the guidelines of the Animal Research Ethics Committee of the Hainan Provincial Education Centre for Ecology and Environment, Hainan Normal University (No. HNECEE-2012-004) and the Experimental Animal Ethics Committee of Guizhou Normal University (No. 2021001).
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Acknowledgments
We are grateful to Yuhan Zhang and Guixia Wan for their assistance with field experiments. We would like to thank the anonymous reviewers for their constructive comments on our manuscript.
Anderson, M.G., Ross, H.A., Brunton, D.H., Hauber, M.E., 2009. Begging call matching between a specialist brood parasite and its host: a comparative approach to detect coevolution. Biol. J. Linn. Soc. 98, 208–216.
Bates, D., Maechler, M., 2009. Package "lme4". . (Accessed 20 December 2023).
Beecher, M.D., Beecher, I.M., Hahn, S., 1981. Parent-offspring recognition in bank swallows (Riparia riparia): Ⅱ. Development and acoustic basis. Anim. Behav. 29, 95–101.
Bioacoustics Research Program, 2014. Raven Pro. Interactive Sound Analysis Software, Version 1.5 Computer Software. The Cornell Lab of Ornithology, Ithaca, NY, USA.
Boncoraglio, G., Saino, N., 2008. Barn swallow chicks beg more loudly when broodmates are unrelated. J. Evol. Biol. 21, 256–262.
Briskie, J.V., Martin, P.R., Martin, T.E., 1999. Nest predation and the evolution of nestling begging calls. Proc. R. Soc. Lond. B Biol. Sci. 266, 2153–2159.
Butchart, S., Kilner, R., Fuisz, T., Davies, N., 2003. Differences in the nestling begging calls of hosts and host-races of the common cuckoo, Cuculus canorus. Anim. Behav. 65, 345–354.
Collar, N., 2020. Daurian redstart (Phoenicurus auroreus). In: del Hoyo, J., Elliott, A., Sargatal, J., Christie, D.A., de Juana, E. (Eds.), Birds of the World, version 1.0. Cornell Lab of Ornithology, Ithaca, NY, USA.
Davies, N.B., 2000. Cuckoos, Cowbirds and Other Cheats. T & AD Poyser. London.
Davies, N.B., 2011. Cuckoo adaptations: trickery and tuning. J. Zool. 284, 1–14.
Davies, N., Madden, J., Butchart, S., Rutila, J., 2006. A host-race of the cuckoo Cuculus canorus with nestlings attuned to the parental alarm calls of the host species. Proc. R. Soc. Lond. B Biol. Sci. 273, 693–699.
Dawkins, R., Krebs, J.R., 1979. Arms races between and within species. Proc. R. Soc. Lond. B Biol. Sci. 205, 489–511.
De Mársico, M.C., Gantchoff, M.G., Reboreda, J.C., 2012. Host–parasite coevolution beyond the nestling stage? Mimicry of host fledglings by the specialist screaming cowbird. Proc. R. Soc. Lond. B Biol. Sci. 279, 3401–3408.
Feeney, W.E., Welbergen, J.A., Langmore, N.E., 2012. The frontline of avian brood parasite–host coevolution. Anim. Behav. 84, 3–12.
Fraga, R.M., 1998. Interactions of the parasitic screaming and shiny cowbirds (Molothrus rufoaxillaris and M. bonariensis) with a shared host, the bay-winged cowbird (M. badius). In: Rothstein, S.I., Robinson, S.K. (Eds.), Parasitic Birds and Their Hosts: Studies in Coevolution. Oxford University Press, Oxford, UK, pp. 173–193.
García, N.C., Barreira, A.S., Kopuchian, C., Tubaro, P.L., 2014. Intraspecific and interspecific vocal variation in three Neotropical cardinalids (Passeriformes: fringillidae) and its relationship with body mass. Emu 114, 129–136.
Glassey, B., Forbes, S., 2002. Muting individual nestlings reduces parental foraging for the brood. Anim. Behav. 63, 779–786.
Gloag, R., Kacelnik, A., 2013. Host manipulation via begging call structure in the brood-parasitic shiny cowbird. Anim. Behav. 86, 101–109.
Grim, T., 2007. Experimental evidence for chick discrimination without recognition in a brood parasite host. Proc. R. Soc. Lond. B Biol. Sci. 274, 373–381.
Haskell, D.G., 1999. The effect of predation on begging-call evolution in nestling wood warblers. Anim. Behav. 57, 893–901.
Honza, M., Vošlajerová, K., Moskát, C., 2007. Eviction behaviour of the common cuckoo Cuculus canorus chicks. J. Avian Biol. 38, 385–389.
Jamie, G.A., Kilner, R.M., 2017. Begging call mimicry by brood parasite nestlings: adaptation, manipulation and development. In: Soler, M. (Ed.), Avian Brood Parasitism: Behaviour, Ecology, Evolution and Coevolution. Springer International Publishing, Cham, pp. 517–538.
Jamie, G.A., Van Belleghem, S.M., Hogan, B.G., Hamama, S., Moya, C., Troscianko, J., et al., 2020. Multimodal mimicry of hosts in a radiation of parasitic finches. Evolution 74, 2526–2538.
Kedar, H., Rodrıguez-Gironés, M., Yedvab, S., Winkler, D., Lotem, A., 2000. Learning modifies offspring signalling of need to their parents. Proc. R. Soc. Lond. B Biol. Sci. 267, 1723–1727.
Kilner, R.M., Madden, J.R., Hauber, M.E., 2004. Brood parasitic cowbirds use host young to procure food. Science 305, 877–879.
Kilner, R., Noble, D., Davies, N.B., 1999. Signals of need in parent–offspring communication and their exploitation by the common cuckoo. Nature 397, 667–672.
Kölliker, M., Brinkhof, M.W., Heeb, P., Fitze, P.S., Richner, H., 2000. The quantitative genetic basis of offspring solicitation and parental response in a passerine bird with biparental care. Proc. R. Soc. Lond. B Biol. Sci. 267, 2127–2132.
Lama, F., Ursino, C.A., Reboreda, J.C., De Mársico, M.C., 2022. Acoustic discrimination by hosts favours vocal trickery in fledglings of the brood-parasitic screaming cowbird. Behav. Ecol. Sociobiol. 76, 71.
Langmore, N.E., Hunt, S., Kilner, R.M., 2003. Escalation of a coevolutionary arms race through host ejection of brood parasitic young. Nature 422, 157–160.
Langmore, N.E., Maurer, G., Adcock, G.J., Kilner, R.M., 2008. Socially acquired host-specific mimicry and the evolution of host races in horsfield's bronze-cuckoo Chalcites basalis. Evolution 62, 1689–1699.
Leonard, M.L., Horn, A.G., 2001. Begging calls and parental feeding decisions in tree swallows (Tachycineta bicolor). Behav. Ecol. Sociobiol. 49, 170–175.
Leonard, M.L., Horn, A.G., 2008. Does ambient noise affect growth and begging call structure in nestling birds? Behav. Ecol. 19, 502–507.
Leonard, M.L., Horn, A.G., Parks, E., 2003. The role of posturing and calling in the begging display of nestling birds. Behav. Ecol. Sociobiol. 54, 188–193.
Liénard, J.-S., Di Benedetto, M.-G., 1999. Effect of vocal effort on spectral properties of vowels. J. Acoust. Soc. Am. 106, 411–422.
Liu, X., Zhang, Y., Wan, G., Luo, H., Wang, L., Liang, W., 2023. First description of injury feigning behaviour in Godlewski's buntings. Ecol. Evol. 13, e10028.
Lotem, A., 1993. Learning to recognize nestling is maladaptive for cuckoo Cuculus canorus hosts. Nature 362, 743–744.
Madden, J.R., Davies, N.B., 2006. A host-race difference in begging calls of nestling cuckoos Cuculus canorus develops through experience and increases host provisioning. Proc. R. Soc. Lond. B Biol. Sci. 273, 2343–2351.
Magrath, R.D., Haff, T.M., Horn, A.G., Leonard, M.L., 2010. Calling in the face of danger: predation risk and acoustic communication by parent birds and their offspring. Adv. Stud. Behav. 41, 187–253.
McLean, I.G., Waas, J.R., 1987. Do cuckoo chicks mimic the begging calls of their hosts? Anim. Behav. 35, 1896–1898.
Mikula, P., Valcu, M., Brumm, H., Bulla, M., Forstmeier, W., Petrusková, T., et al., 2021. A global analysis of song frequency in passerines provides no support for the acoustic adaptation hypothesis but suggests a role for sexual selection. Ecol. Lett. 24, 477–486.
Noh, H.-J., Gloag, R., Leitão, A.V., Langmore, N.E., 2021. Imperfect mimicry of host begging calls by a brood parasitic cuckoo: a cue for nestling rejection by hosts? Curr. Zool. 67, 665–674.
R Development Core Team R, 2018. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria.
Redondo, T., Reyna, L.A.D., 1988. Vocal mimicry of hosts by great spotted cuckoo clamator glandarius: further evidence. Ibis 130, 540–544.
Rojas Ripari, J.M., Ursino, C.A., Reboreda, J.C., De Mársico, M.C., 2019. Innate development of acoustic signals for host parent–offspring recognition in the brood-parasitic screaming cowbird Molothrus rufoaxillaris. Ibis 161, 717–729.
Rojas Ripari, J.M., Ursino, C.A., Reboreda, J.C., De Mársico, M.C., 2021. Tricking parents: a review of mechanisms and signals of host manipulation by brood-parasitic young. Front. Ecol. Evol. 9, 725792.
Roldán, M., Martín-Gálvez, D., Rodríguez, J., Soler, M., 2013. Breeding biology and fledgling survival in a carrion crow Corvus corone population of southern Spain: a comparison of group and pair breeder. Acta Ornithol. 48, 221–235.
Ruiz-Raya, F., Soler, M., Soler, M., 2017. Avian Brood Parasitism – Behaviour, Ecology, Evolution and Coevolution. Springer International Publishing AG, Cham, Switzerland.
Samaš, P., Žabková, K., Petrusková, T., Procházka, P., Požgayová, M., Honza, M., 2020. Nestlings of the common cuckoo do not mimic begging calls of two closely related Acrocephalus hosts. Anim. Behav. 161, 89–94.
Sato, N.J., Tokue, K., Noske, R.A., Mikami, O.K., Ueda, K., 2010. Evicting cuckoo nestlings from the nest: a new anti-parasitism behaviour. Biol. Lett. 6, 67–69.
Schuetz, J.G., 2005. Reduced growth but not survival of chicks with altered gape patterns: implications for the evolution of nestling similarity in a parasitic finch. Anim. Behav. 70, 839–848.
Soler, M., 2017. Begging behaviour, food delivery and food acquisition in nests with brood parasitic nestlings. In: Soler, M. (Ed.), Avian Brood Parasitism: Behaviour, Ecology, Evolution and Coevolution. Springer International Publishing, Cham, pp. 493–515.
Tokue, K., Ueda, K., 2010. Mangrove gerygones gerygone laevigaster eject little bronze-cuckoo Chalcites minutillus hatchlings from parasitized nests. Ibis 152, 835–839.
Tuero, D.T., Gloag, R., Reboreda, J.C., 2016. Nest environment modulates begging behavior of a generalist brood parasite. Behav. Ecol. 27, 204–210.
Ursino, C.A., Gloag, R., Reboreda, J.C., De Mársico, M.C., 2018. Host provisioning behavior favors mimetic begging calls in a brood-parasitic cowbird. Behav. Ecol. 29, 328–332.
Wan, G., Zhao, H., Liu, X., Wang, L., Liang, W., 2023. Predation of Daurian redstarts offspring in nest boxes by the Oriental magpie-robin and tree sparrow. Ecol. Evol. 13, e10093.
Wang, J., Li, Q., Yang, C., 2020. Coevolution of acoustical communication between obligate avian brood parasites and their hosts. Avian Res. 11, 43.
West, M.J., King, A.P., 1988. Female visual displays affect the development of male song in the cowbird. Nature 334, 244–246.
Yang, C., Liang, W., Antonov, A., Cai, Y., Stokke, B.G., Fossøy, F., Moksnes, A., Røskaft, E., 2012. Diversity of parasitic cuckoos and their hosts in China. Chin. Birds 3, 9–32.
Yang, C., Si, X., Liang, W., Møller, A.P., 2020. Spatial variation in egg polymorphism among cuckoo hosts across 4 continents. Curr. Zool. 66, 477–483.
Yang, C., Wang, L., Cheng, S.-J., Hsu, Y.-C., Liang, W., Møller, A.P., 2014. Nest defenses and egg recognition of yellow-bellied prinia against cuckoo parasitism. Naturwissenschaften 101, 727–734.
Zheng, G., 2023. A Checklist on the Classification and Distribution of the Birds of China, fourth ed. Science Press, Beijing.
Zhong, G., Wan, G., Zhang, Y., Zhao, H., Wang, L., Liang, W., 2023. Nest desertion as an anti-parasitism strategy in hosts selects for late egg-laying behavior in cuckoos. iScience 26, 108156.
Table
1.
Analysis of variance of age matched begging call features of cuckoo nestlings and host species' nestlings raised in four host species' nests (SE refers to standard error and significant results are shown in bold).
Table
2.
Linear regression analysis of begging calls of cuckoo nestlings and four host species nestlings against age (SE refers to standard error and significant results are shown in bold).

 DownLoad:
DownLoad:





 Email Alerts
Email Alerts RSS Feeds
RSS Feeds