Canchao YANG, Yan CAI, Wei LIANG. 2012: Species identification of sympatric cuckoo nestlings in a multiple-cuckoo system, China. Avian Research, 3(2): 108-112. DOI: 10.5122/cbirds.2012.0016
Citation:
Canchao YANG, Yan CAI, Wei LIANG. 2012: Species identification of sympatric cuckoo nestlings in a multiple-cuckoo system, China. Avian Research, 3(2): 108-112. DOI: 10.5122/cbirds.2012.0016
Canchao YANG, Yan CAI, Wei LIANG. 2012: Species identification of sympatric cuckoo nestlings in a multiple-cuckoo system, China. Avian Research, 3(2): 108-112. DOI: 10.5122/cbirds.2012.0016
Citation:
Canchao YANG, Yan CAI, Wei LIANG. 2012: Species identification of sympatric cuckoo nestlings in a multiple-cuckoo system, China. Avian Research, 3(2): 108-112. DOI: 10.5122/cbirds.2012.0016
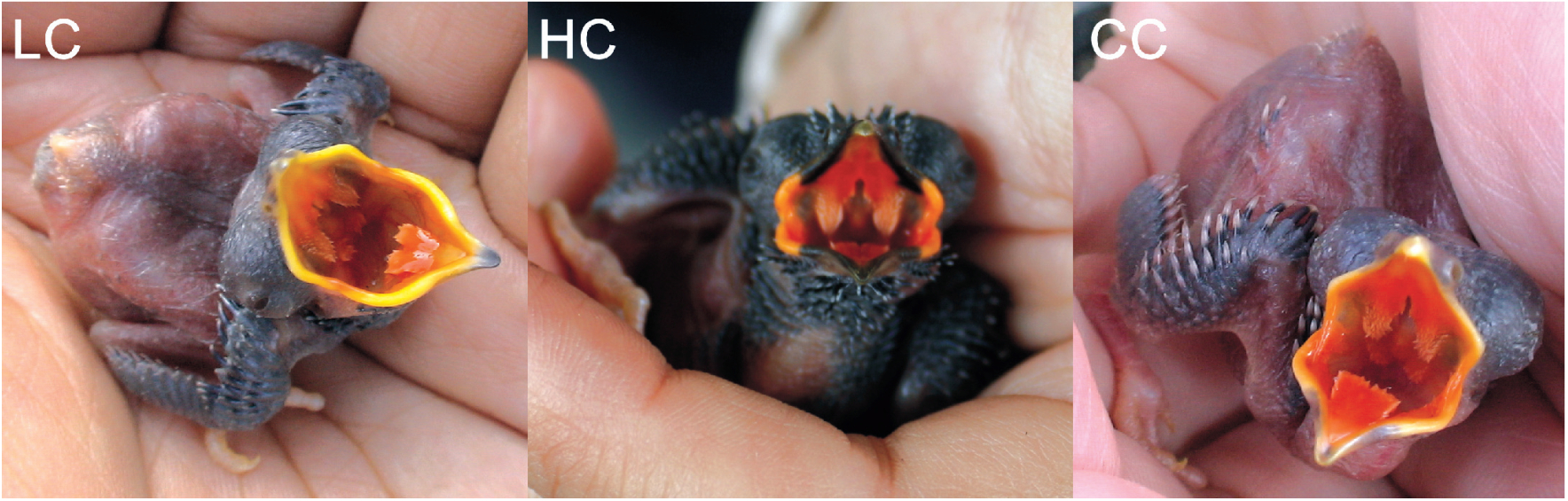
Interaction between a parasite and its host could lead to a co-evolutionary arms race. Cuckoo-host system is among the most studied of all brood parasite systems, but the cuckoos of Asia, on the other hand, are much less well known. China has the most abundant cuckoo species in Asia. Many of these co-occur in sympatric areas, posing a potential risk of mis-identification of cuckoo nestlings, especially in Cuculus species. In this study we have provided a practical criterion to identify cuckoo nestlings species in the field and performed molecular phylogeny to confirm our empirical results. These results indicate that two distinct characteristics of cuckoo nestlings, i.e., the gape color pattern and feather traits can be considered as reliable species identification. To our knowledge, this is the first report for species identification of Cuculus nestlings through molecular analysis.
Our current understanding is that the Helmeted Guineafowl flock is basically a group of birds that remain together for at least one breeding season and that members follow a dominant male during foraging in the non-breeding season (dry season, May to September in the Krugersdorp Game Reserve). One function of this behavior is to maximize the discovery of underground food resources that occur in patches (Van Niekerk, 1979, 1980, 2002; Crowe and Elbin, 1987). Ecological constraints are lifted as rain falls (October in the Krugersdorp Game Reserve) and the members engage in sequential mate selection activities. Monogamous pairs form and males protect their females and chicks (Crowe and Elbin, 1987). According to Elbin (1979) the prime reproductive system is the family unit (mother, father and offspring) as revealed by studies conducted on free-ranging Domestic Guineafowl in Pennsylvania (USA). In short, behavioral studies have concentrated on seasonal events and the reproductive system of Helmeted Guineafowl (Skead, 1962; Elbin, 1979), while this paper focuses on the behavioral patterns of individuals and how they contribute to the maintenance of the flock. More specifically the purposes of this paper are: 1) to describe the cyclic re-grouping of members within the flock to maintain cohesion and flock stability and 2) to describe patterns of co-operation within the flock that have not been the focus of previous studies.
Study area and methods
This study was conducted in the 1400-ha Krugersdorp Game Reserve (KGR) (26°05′11″S, 27°42′35″E), in Gauteng Province, South Africa. From March to July 1981, seven flocks (8–15 members) in a designated area were trapped using walk-in funnel traps. A total of 52 Helmeted Guineafowl were fitted with metal rings and numbered patagial tags, representing at least 40% of all members in each flock (Van Niekerk, 1983, 2009). Adults were sexed through cloacal examination (Siopes and Wilson, 1973). These flocks were visited weekly from April 1982 to March 1983 and then monthly from April 1983 to February 1984. With the aid of a mounted telescope all marked guineafowl were noted on standard maps while the author traversed a fixed route during late afternoons, totaling ~150 hours of observations.
Periods of 45–60 min were spent in close proximity to one flock that was selected randomly for intensive study during each visit from April 1982 to March 1983, totaling about 60 hours of observations (Van Niekerk, 2009). In late afternoons the study flock was mainly located in a small area (400 m2), which made it possible to place permanent beacons that would allow for the estimation of distances between members. The association time between individuals and all interactive behavioral patterns (such as pecking) were recorded. Individuals were regarded as associated when they foraged close to one another (30–100 cm apart with no other guineafowl between them) for 60 s or more. Social spread (m) is the direct distance between the two furthest members through the flock (excluding breeding pairs). Where information was lacking, facts were gleaned from a similar unpublished study of a flock of 10 Helmeted Guineafowl that visited a private garden near Constantia (33°59′45″S, 18°26′06″E) in Cape Town. This study (January 1992 to May 1994) was done by Ereka van der Merwe, who distinguished individuals on the basis of casque size and shape (Prinsloo et al., 2005). Females and males were distinguished on the basis of fights (only males engage in fights) and calls (only females emit a "buckwheat" call). Agonistic displays refer to chases, pecks, humpbacked postures and head-bobbing by high ranking males in the direction of a low ranking member or incoming conspecific intruder.
Results
Cyclic events
During the breeding season (October–March) the seven flocks remained in small separate breeding territories (0.5–1 ha), but joined neighboring flocks during the non-breeding season (April–September) to form larger flocks (~30 individuals) that foraged across a larger area (71 ha). Juveniles from the different flocks were pooled from April onwards, which was a period when flocks moved further away from cover as their offspring grew from chicks to the juvenile stage (Siegfried, 1966). This was necessary since food resources had been depleted (Van Niekerk, 2002; Van Niekerk, unpublished). Despite the apparent joining of neighboring flocks, the marked flocks returned to their respective roosts during the late afternoon. In fact, the colored patagial tags revealed that the separate flocks remained in their respective subgroups even while they were with the larger congregated flock.
Flock size and structure
The mean monthly size of all flocks in the study was 7.75 (SD = 3.02, n = 33, range = 7–14) (Van Niekerk, 1985). The study flock basically consisted of three adult males (A, B and C), three adult females (D, E and F), four to six offspring and two non-permanent females that sometimes visited the flock for two or three days at a time. At least two other flocks had two or more adult males and females, but during the 1981/82 breeding season not one marked flock succeeded in raising more than one brood of chicks.
Social ranking and spatial position of members in the study flock
Male A was the highest ranking male because 1) he was at the top of the pecking order (Table 1), 2) he initiated foraging direction and the rest of the flock followed him, 3) he was the first to court and breed in two consecutive breeding seasons and 4) he was the member that initiated action to chase intruding guineafowl from other flocks that had moved into the breeding territory (Van Niekerk, 2002; Van Niekerk, unpublished). During the non-breeding season (April–October), the other two males remained close to him, with females and offspring further behind. The non-breeding male often ran towards male A and performed a submissive display in front of male A (Van Niekerk, 1985).
Table
1.
Number of agonistic interactions of members of the study flock arranged in a sociogram
Females usually remained between the adult males and their offspring. No rank order was observed among females. The non-breeding female was often observed in a tree above the flock from where she emitted a "buckwheat" call, which is a typical female call (Maier, 1982).
Close-knit unit
The study flock remained a permanent group and members were often found close together throughout the study period. The social spread of the flock was 25–40 m during the period before the chicks arrived and 5–10 m just after their arrival (Fig. 1). In fact, there was a significant negative correlation between social spread and mean flock size (r = –0.79, p < 0.01). The shorter distance during the brooding phase (January–February) was a direct consequence of clustering around the chicks (1–10 days old). The chicks moved along with the flock, often directly beneath the adults and made body contact with various adults that emitted a soft "swe swe swe" contact call. When approached by humans (n = 10), the Black-backed Jackal (Canis mesomelas) (n = 6) and the Savanna Baboon (Papio ursinus) (n = 8), some adult members scattered in different directions while the entire flock produced an inferno of panic calls to distract the intruders (Maier, 1982). The chicks also scattered and hid in nearby grass. As the chicks reached juvenile age, they formed a separate subgroup right behind the flock, about 1–3 m away. This distance increased as they reached sub-adulthood during the non-breeding season (April–September). This was the time when a troop of baboons was seen killing one sub-adult and probably more, as the number of sub-adults dwindled from six to three during the first year. The 1981/82 offspring remained with the flock for at least 18 months.
Figure
1.
Social spread correlated with mean group sizes of the study flock based on four observations per month
The study flock did not dissolve when male A and female D paired off to breed nearby for a period of ~28 days during December 1981, who remained only about 20 m from the area where the rest of the flock usually foraged. In fact, the remainder of the flock continued to forage in a territory of ~0.5 ha around the nest. Male A was not regularly observed in the flock during incubation, but did arrive from time to time without his female, who was incubating their eggs. During the 1981/82 breeding season male B and female E paired off to produce chicks after A and D returned to the flock with their chicks in January 1982. However, this second brood perished within one month.
A comparison of the agonistic displays by the two high ranking males (A and B) towards members or intruders and low ranking members (such as humpbacked displays and chases) demonstrates that B was less agonistic than A in the presence of A. Once A paired off to breed and moved further away from the flock, B showed more agonistic displays and assumed the role of dominant male. The rest of the flock clustered around him during the absence of A. The position of A was restored once he returned to the flock in March, when both males were present together again (Fig. 2).
Figure
2.
Alternate frequencies of agonistic displays by the two highest ranking males in the study flock based on 45 hours of observations, totaling 62 displays
The study flock remained together during at least two breeding seasons up to March 1983. The chicks of the second breeding season joined the offspring from the previous year in a subgroup. Female D disappeared during March 1982, but was replaced by a new female during the following breeding season. The Constantia group showed a similar degree of stability over a two-year period.
Associations within the study flock
Table 2 demonstrates that adult males clustered together just prior to and during the first part of the breeding season. In fact, when intruding conspecifics approached these males stood together side by side and moved together in the direction of the imposters. These bonds became less prevalent during the period from the arrival of the chicks to the non-breeding season (Table 3), when the high ranking males spent more time with the chicks (Table 4). Breeding females (D and E) spent more time with their own males than with the non-breeding male (respectively χ2 = 656.7, df = 2, p < 0.01 and χ2 = 1201.1, df = 2, p < 0.01; Table 3). This is expected in a monogamous mating system. However, the second ranking male (male B) spent more time with his female than the highest ranking male (male A) did during flocking (Table 3). The non-breeding male spent most of his time with the higher ranking males (A and B) and to a less extent with the breeding females (D and E) (χ2 = 502.47, df = 3, p < 0.01), but did not engage in any courtship behavior (Table 3). Similarly, the non-breeding female (F) spent most of her time with the higher ranking males and also did not perform any courtship behavior. Her association with the breeding females (D and E) was virtually non-existent (Tables 3) (χ2 = 631.6, df = 3, p < 0.01).
Table
2.
Association patterns of adult members of the study flock in % and time (min) from September 1982 to January 1983
Table
3.
Association patterns of adult members of the study flock in % and time (min) from February 1983 to May 1983 while observed in the flock (pairs were absent from time to time)
Table 4 shows the time flock members spent with male A's chicks. The mother (D) was the least caring (Table 4). The father (A) and the second ranking male (B) spent the most time with the chicks. The non-breeding male and female spent more time with the chicks than the breeding females (χ2 = 205.8, df = 5, p < 0.01). In the Constantia flock, an adult male member, which was not the father, often spread his wings over the chicks to shield them from intruders such as dogs and humans. In the same flock, the off-spring of the breeding female from the previous year helped with incubation when the mother got up to feed.
Leadership succession
Male A remained the highest ranking male for at least two breeding seasons and produced chicks during both. He was never challenged (involved in physical battles) by a male within the flock. However, in the Constantia flock, a new male and female joined the original flock from the nearby Kirstenbosch Botanical Gardens. When they joined, a bloody physical battle that lasted for days ensued between the new male and the original highest ranking male. Blood oozed from their bare facial parts as they pecked at each other's wattles. The new male did not succeed and remained on the periphery of the flock during the next 10 months. During the start of the next breeding season, the new male launched a renewed attack on the highest ranking male and succeeded. The old male and his female retired to the side of the flock while the new male started initiating foraging direction with the rest of the flock following.
Discussion and conclusions
The study flock was not a number of Helmeted Guineafowl that associated randomly. The members remained around the dominant male and followed him as he initiated foraging. Although a pecking order could be established among males, the frequency of agonistic actions was very low, which suggests that agonistic interactions are limited to ensure cohesion. The function of the pecking order was not to dispel adult members from the flock but simply to maintain a rank hierarchy. The second highest ranking male took center stage with the rest of the flock associated around him in order to maintain the cohesive nature of the flock when the dominant male and his female left to breed. This alternate responsibility was important to prevent eviction from the territory or a take-over from neighboring dominant males. The two high ranking males foraged away from the females during the non-breeding season, but still in the same flock, in order to repel conspecific intruders, but during the breeding season they spent more time with their females (Table 3). In other words, it was a monogamous mating system. No polygamy or promiscuity was recorded; that is, the two high ranking males did not flirt with each other's females or flirt with females that joined from neighboring flocks. Prolonged squabbles for adult females could lead to flock instability. There was no evidence of sequential mate selection. Flirting by paired males was recorded by Elbin et al. (1986) elsewhere.
On arrival of parents and chicks, the highest ranking male spent more time with his chicks than the mother, who spent relatively little time with her chicks (Table 4). This is ascribed to the females foraging actively to restore bodily condition after breeding (Crowe and Elbin, 1987). During this time, the adult non-breeding male and female spent more time with the chicks of the dominant male than their own mother did (Table 4). Since the history of the flock was not known, these helpers could be offspring from a previous year since they were at least two years old when observations started. In the Constantia study, it was demonstrated that offspring from a previous year helped their mother with incubation.
The small social spread during the brooding stage illustrates that most members form a close-knit unit to protect the chicks. It could be argued that formation of a cluster is a response to an urge to associate closely when predators close in, because chicks are easy prey (Fig. 1). However, the fact that all adults 1) helped to distract predators, 2) emitted soft "swe swe swe" contact calls and 3) associated closely with the chicks, illustrates that co-operative nurturing indeed took place.
Unlike free ranging Domestic Guineafowl, where the family is the primary reproductive and brooding unit (Elbin, 1979), this study shows that wild Helmeted Guineafowl base their survival on co-operation in a flock with a number of adult Helmeted Guineafowl. Some adults did not pair off to breed, whereas breeding appears to be the prerogative of the highest ranking male, followed by the second highest ranking male. However, intrinsic/extrinsic triggers could determine how many guineafowl pairs form from a flock (Van Niekerk, unpublished). Extrinsic factors could be predator pressure and rainfall, while intrinsic factors could include how many offspring remained in the flock from the previous year (cf. Crowe, 1978b).
Among Crested Dendroperdix sephaena and Coqui Peliperdix coqui Francolins, the mother and father produce a brood and when their offspring reach sub-adulthood (7–8 months old) they are repelled from the covey (Van Niekerk, 2001; Van Niekerk and Van Ginkel, 2003). Among Red-necked Pternistis afer and Swainson's P. swainsonii Spurfowls, the father occupies a territory and adult females that enter are courted and inseminated. The mother raises her chicks without the assistance of the father, but the chicks are also repelled when they are about seven to eight months old (Van Niekerk et al., 2009; Van Niekerk, unpublished). In comparison, Helmeted Guineafowl live in flocks and their offspring remain in the flock for at least 18 months. Pairs break away from the flock to breed and return to the flock with their chicks. The non-breeding flock members assist in raising the chicks.
Although Helmeted Guineafowl live in flocks, the breeding pairs actually fare less well than Coqui or Crested Francolin pairs in terms of reproductive efficiency. Efficiency is the mean number of juveniles from a brood expressed as a percentage of the mean clutch size. The mean clutch sizes are 12 for Helmeted Guineafowl, 5.8 for Swainson's Spurfowl, 4.8 for Crested Francolin and 5.0 for Coqui Francolin (Hockey et al., 2005). Reproductive efficiency is 38.2% for Helmeted Guineafowl, 36.6% for Swainson's Spurfowl, 41.8% for Crested Francolin and 44.0% for Coqui Francolin: χ2 = 45.5, df = 3, p < 0.01 (Van Niekerk, unpublished).
Clearly, if the flock is compared to the family system of the francolins, it is not necessarily advantageous for a higher juvenile output. In fact, flocking as a protective unit strategy becomes less convincing if one considers that in the case of the francolins only two adults look after their offspring, while in the case of spurfowls only the mother raises the chicks. However, unlike the francolins and spurfowls, Helmeted Guineafowl move widely in open country where the vigilance potential of more members minimizes surprise attacks and so enhances chick survival (Treisman, 1975). Secondly, flocking is an effective strategy to repulse conspecific intruders from encroaching on their established territory. This last point became especially evident when the non-breeding members remained in the territory and protected it during the absence of the highest ranking male.
To summarize, cooperation within the flock was revealed by: 1) males that joined to repel intruders, 2) the second highest ranking male taking center stage in the flock while the highest ranking male was absent, 3) shared brooding functions among adult members and 4) predator evasion, especially when chicks were nurtured jointly.
Davies NB. 2000. Cuckoos, Cowbirds and Other Cheats. Poyser, London.
Payne RB. 2005. The Cuckoos. Oxford University Press, New York.
Rothstein SI, Robinson SK. 1998. Parasitic Birds and Their Hosts. Oxford University Press, New York.
Yang C, Antonov A, Cai Y, Stokke BG, Moksnes A, Røskaft E, Liang W. 2012b. Large Hawk-cuckoo Hierococcyx sparverioides parasitism on the Chinese Babax Babax lanceolatus may be an evolutionarily recent host-parasite system. Ibis, 154: 200–204.
Yang C, Cai Y, Liang W. 2011. Visual modeling reveals cryptic aspect in egg mimicry of Himalayan Cuckoo (Cuculus saturatus) on its host Blyth's Leaf Warbler (Phylloscopus reguloides). Zool Res, 32: 451–455.
Yang C, Cai Y, Liang W. 2010b. Brood parasitism and egg mimicry on Brownish-flanked Bush Warbler (Cettia fortipes) by Lesser Cuckoo (Cuculus poliocephalus). Zool Res, 31: 555–560. (in Chinese)
Yang C, Liang W, Antonov A, Cai Y, Stokke BG, Fossøy F, Moksnes A, Røskaft E. 2012a. The diversity of parasitic cuckoos and their hosts in China. Chinese Birds, 3: 9–32.
Yang C, Liang W, Cai Y, Shi S, Takasu F, Møller AP, Antonov A, Fossøy F, Moksnes A, Røskaft E, Stokke BG. 2010a. Coevolution in action: disruptive selection on egg colour in an avian brood parasite and its host. PLoS ONE, 5(5): e10816.
Zheng G. 2011. A Checklist on the Classification and Distribution of the Birds of China. 2nd edn. Science Press, Beijing.
Table
3.
Association patterns of adult members of the study flock in % and time (min) from February 1983 to May 1983 while observed in the flock (pairs were absent from time to time)

 DownLoad:
DownLoad:







 Email Alerts
Email Alerts RSS Feeds
RSS Feeds