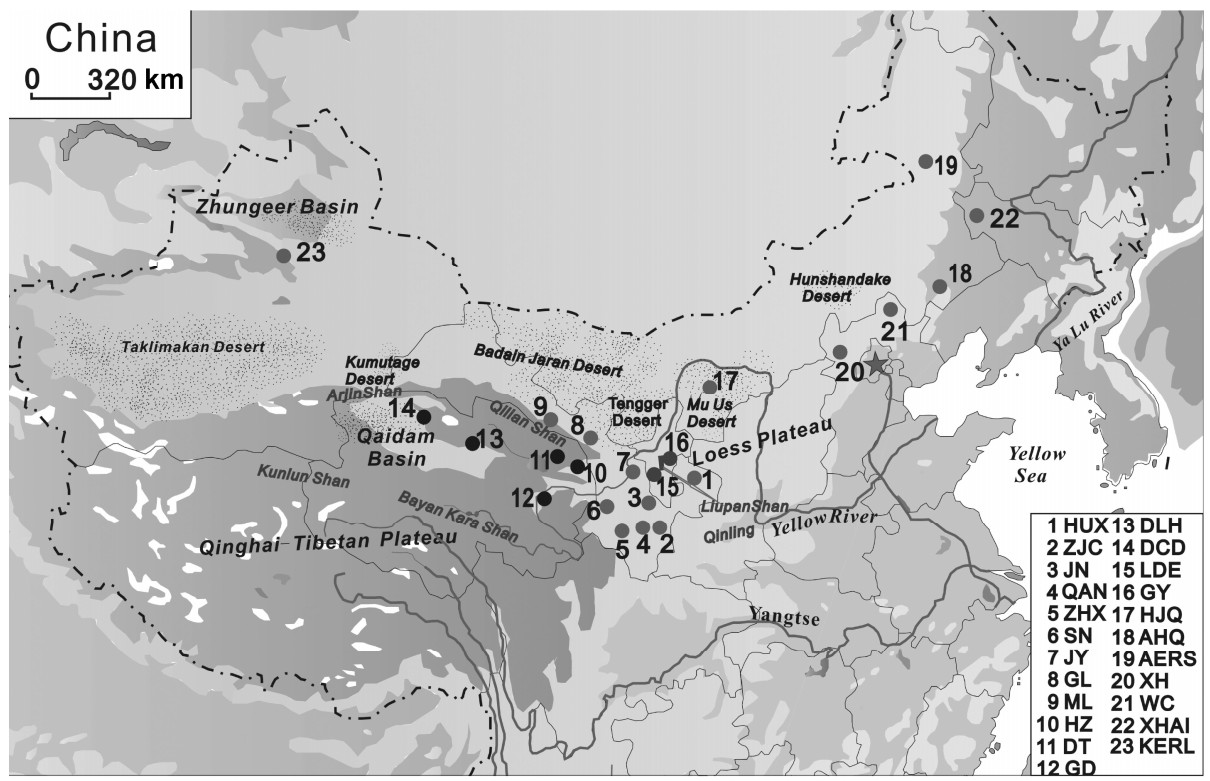
The Daurian Partridge (Perdix dauuricae) is a kind of hunting bird with high economic value. Genetic diversity and structure in the Daurian Partridge were studied by analyzing eight microsatellite loci in 23 populations found throughout the range of the species in China. The objectives were to evaluate the consequences on genetic diversity and differentiation of Daurian Partridge populations and to obtain a profound genetic insight for future management decisions and for effective measures to protect and exploit Daurian Partridges. The results showed that microsatellites were polymorphic in all Daurian Partridge populations, with a high level of genetic diversity over all the loci, especially in the Qaidam Basin populations which have the highest level of diversity. Significant genetic divergence was observed among different groups as well as between populations within the same group; most pairwise FST values were highly significant. Both phylogenetic trees and Bayesian clustering analyses revealed clear differentiation among the 23 populations of the Daurian Partridge, which were classified into two genetically differentiated groups. A bottleneck analysis indicated that Daurian Partridge populations have experienced a recent bottleneck. Our study argues that the Qaidam populations, North China populations, JN population, ZJC population, and Liupan Mountain populations should be paid special attention in order to retain adequate population sizes for maintaining genetic diversity.
Waterbird populations are declining worldwide, especially in Asia (Wetlands International, 2006), with the situation particularly critical in eastern China where the fast rate of economic development has caused high levels of land reclamation, pollution and human disturbance of both coastal and inland wetlands (NWCAPC, 2000; He and Zhang, 2001; Gong et al., 2010) which, historically, have supported large numbers of wintering waterbirds with a high level of diversity (Lu, 1996; Kear, 2005).
The Yangtze River floodplain is the most important area for waterbirds in eastern China, with extensive waterbird counts conducted during the 2004 and 2005 winters, indicating that the floodplain supports more than one million waterbirds during the non-breeding period (Barter et al., 2004, 2006). However, it has been estimated that the number of Anatidae birds in eastern China have declined by about 70% in the last 20 years (Cao et al., 2008a); populations of other waterbird species are also likely to have suffered a similar decline (e.g. Cao et al., 2008b; Shi et al., 2008). Threats to the wetlands of the Yangtze River floodplain will continue to intensify as massive hydrological changes occur (BirdLife International, 2003; Zhao et al., 2005; Yang et al., 2006, 2007; Cao and Fox, 2009). In addition, it is predicted that climate change will cause 85% of global inland wetlands to be lost by the end of this century (UNEP, 2009) and it is likely that losses in China will be even greater, for the temperatures in eastern China are expected to rise even more than the global average (Anon, 2007). The potential for impending changes in habitat areas and their quality are further expected to have a negative impact on the waterbirds of the Yangtze River floodplain, which makes it imperative to monitor trends in the number of waterbirds and their distribution in the region, so that effective science-based conservation measures can be initiated to ensure their survival and future recovery.
Obtaining accurate data on the number of waterbirds and their distribution, which can be confidently compared across years and sites, is complicated by a number of factors which can affect the quality of these counts, such as 1) different survey methods, 2) variation in observer experience, 3) disparities in environmental conditions and 4) changes in human disturbance levels. Thus, it is essential to employ a survey methodology which maximizes count accuracy and precision and minimizes the biases inherent in counting waterbirds (Erwin, 1982; Howe et al., 2000; Atkinson et al., 2006; Rogers et al., 2006; Haslem et al., 2008). Failure to achieve these goals will lead to incorrect results and analytical problems, such as the inability to assess the statistical significance of apparent trends (Thomas, 1996; Thomas and Martin, 1996; Gosbell and Clemens, 2006).
Recently we developed a systematic waterbird survey methodology which was tested with promising results during the 2008/2009 and 2009/2010 winter seasons at Shengjin Lake, an important wetland in the Yangtze River floodplain. The methodology draws on recent improvements to an extensive Australian shorebird count program which has operated since 1980 (Haslem et al., 2008). Analysis of the first 28 years of this data showed that the existing shorebird count data set had limited value for detecting population trends and determining their causes. In order to increase data utility, important additions have been made to the types of data collected, improvements were made to data acquisition methods and the sample of shorebird sites has been increased to improve the power to detect population trends; another important objective of the updated Australian program was to investigate the relationship between the distribution and abundance of shorebirds, habitat characteristics and threatening processes (Haslem et al., 2008).
In this paper we provide a detailed description of the survey methodology developed for Shengjin Lake and how it was implemented. We use the results from the first two winters in which the methodology was employed to provide examples of how the data can be used to provide detailed information on the waterbirds and the habitats of Shengjin Lake. Finally, we discuss the importance of wider application of the methodology throughout the Yangtze River floodplain, particularly to provide a robust underpinning to a much needed floodplain-wide waterbird monitoring program.
Methods
Study site
Shengjin Lake (30°16′–30°25′N, 116°59′–117°12′E), part of the Anhui Shengjin Lake National Nature Reserve, is located on the southern bank of the Yangtze River, close to Anqing (Fig. 1). The maximum lake area during the flood peak is ~14000 ha (water level = 17.0 m above sea level (asl); Wusong datum), but the water level normally falls each year to less than 10 m asl during November to February (dry season), causing the lake area to decrease to ~3400 ha. Water comes from three smaller rivers flowing directly into the lake and from the Yangtze River via the Huangpen Sluice (Cheng and Xu, 2005). The average annual rainfall is 1600 mm, with most falling from March to August; average annual temperature is 16.1℃, with an average January temperature of 4.0℃.
Figure
1.
Shengjin Lake (Upper and Lower Lakes), showing location of the Huangpen Sluice which connects the lake to the Yangtze River and survey areas A–E; the inset shows the Yangtze River floodplain with locations of Shengjin, Dongting and Poyang lakes (two other important waterbird wetlands) and the Three Gorges Dam (adapted from Fig. 1 in Cheng et al., 2009).
Shengjin Lake is a particularly important wetland, supporting 5–10% of the total waterbird population within the Yangtze River floodplain (Barter et al., 2004, 2006). The lake is an excellent site for testing the development of a systematic survey methodology because it supports a large and diverse waterbird population spread over a wide variety of habitats subject to all the common threats found in the floodplain. It is also a relatively small wetland that can be satisfactorily surveyed in 2–3 days by small teams of 2–3 people.
Survey methodology
1) The lake was divided into five discrete survey areas (A to E) (Fig. 1). While the major factor taken into account in deciding the size of a survey area was that it had clear geographic boundaries and could be adequately surveyed by a team of 2–3 people in 1/2 to one day, the survey areas also had features that made them distinctive, e.g. areas A and E contained large areas of grazing meadows, B and C were the most highly disturbed regions within the lake, C also had a large number of small bays, and D was a large area of open water important for roosting ducks.
2) Discrete sub-areas were identified within each survey area, each with clear geographic (e.g. shoreline) and artificial (e.g. fishnets, small dams) boundaries which enabled sub-areas to be completely surveyed as distinctive parts of the survey area (Fig. 2 shows how this was achieved for survey area A). There are a number of significant benefits to collecting data on a smaller sub-area scale: i) conducting separate surveys of a number of smaller areas is logistically easier than surveying the complete survey area as one entity; ii) simple corrections can be made for incomplete or varying coverage between surveys; iii) unambiguous identification is available of a distinct region within a larger survey area impacted by a threat and iv) facilitation of habitat analysis, e.g. the effect of habitat type on the number of species and the effect of habitat changes on the number of waterbirds.
Figure
2.
Survey area A showing sub-areas (a1–a8), survey sites (1–11) and GPS route to follow between survey sites, with turnoffs (T1–T5). The survey commences at survey site 1 and continues in an anti-clockwise direction finishing at survey site 11, with a detour to survey sites 4, 5 and 6.
3) Instructions were provided to surveyors describing how to reach each survey site and the sub-area (or part of a sub-area) to be surveyed and a pre-prepared GPS route was supplied to facilitate finding turnoffs and survey sites (Appendix 1 contains the instructions for survey area A).
4) Survey data were recorded in a standardized manner using a separate site survey form for each survey site (i.e. wetland identification, environmental conditions, observer details, number of species, human disturbance and habitat changes: see Appendix 2); instructions were provided on how to complete the site survey form (Appendix 3). Locations of major waterbird concentrations were recorded on maps of the survey areas (see Fig. 2 for example).
5) Photographs were taken, as necessary, of the different habitats and threats to provide a visual record of the conditions existing during the surveys.
Data collection
Survey procedure, frequency and conditions
The general survey procedure involved covering the complete lake in 2–3 days (preferably two, to minimize potential problems with waterbird movements during the survey period). Two teams were used: survey areas A and B were surveyed on the same day by one team while survey area C was being surveyed at the same time by the second team. Survey areas D and E were covered by two teams simultaneously, one surveying the west side and the other the east side of the lake; cell phone communication was used to avoid double counting.
We developed, tested and improved the survey methodology during the 2008/2009 winter, starting in late-November; further improvements were made during the 2009/2010 winter. Seven surveys were conducted in the winter of 2008/2009 (6–8 December; 15–17 December; 5–6 February; 26–27 February; 15–16 March; 30–31 March; 15–16 April) and 13 surveys in the 2009/2010 winter season (11–12 October; 31 October–2 November; 14–15, 18, 21 November; 4–5 December; 18–20 December; 4–6 January; 21–23 January; 4–5, 10 February; 16–17, 19 February; 4–7 March; 16–17 March; 30 March, 3–4 April; 10, 12, 15 April). While all surveys in the winter of 2008/2009 were conducted within the preferred time period (5 × 2 d; 2 × 3 d), weather conditions during six surveys of the 2009/2010 winter surveys were poor (due to continuous foggy conditions, snow, rain, etc.). These were completed over a period of more than three days (extending to a maximum period of eight days).
In 2008/2009, the teams in the first four surveys were led by L Cao and M Barter, each team containing 1–3 students receiving training, while the final three surveys were conducted by the students themselves. All surveys in 2009/2010 were conducted by students, except for partial assistance by L Cao and M Barter on two surveys.
Technique of waterbird counting
We used the "look-see" counting method (Bibby et al., 2000; Delaney, 2005) which is commonly used to count waterbirds (e.g. Banks et al., 2006). The objective of the surveys was to identify and count all waterbirds present in each sub-area, a task made simple by the propensity for most waterbird species to gather in highly visible flocks making them relatively easy to locate and count. However, the less common and cryptic species may be under-counted, especially over extensive areas where they can be hidden by vegetation (Rappoldt et al., 1985).
When counting a species within a sub-area, we recorded the sizes of the individual sub-flocks (component counts) for this reduces the random error associated with counting large numbers of birds (Box 1).
Lake water levels and rainfall
Daily water levels (Wu Song datum) at the Huangpen Sluice (Shengjin Lake and Yangtze River sides) were obtained from the Water Resources Commission. Rainfall data were collected at the Yang'etou Management Station (located on the central east side of the Upper Lake) using a rainfall gauge.
Data treatment and analysis
Box 1 Use of component counts to improve count accuracy
In a large study, consisting of a number of counting experiments in which observers of varying experience counted the same waterbird flocks in a variety of habitats and in which their results were compared with accurate counts of the birds actually present, it was shown that the coefficient of variation (standard deviation (SD)/x) due to observer error and site-specific differences (visibility, distance to birds, vegetation, topography) was 37% (Rappoldt et al., 1985). Since these errors are random they tend to neutralize one another when the counts of a number of individual flocks are combined (Rappoldt et al., 1985). Take, for example, a flock of 761 Bean Geese (Anser fabalis) comprising 10 sub-flocks of 5, 180, 70, 25, 150, 29, 72, 8, 189 and 33. If the count was recorded simply as 761 birds, the SD would be 282 birds (i.e. 761 × 0.37), i.e. there is a 95% certainty that the actual number of birds present was in the range 761 ± 553 (i.e. 1.96 × SD). However, if the count was recorded as 10 separate flocks the SD would be reduced to 119 birds, i.e. there is a 95% certainty that the actual number of birds present was in the range 761 ± 233. Thus, by using component counts a more accurate estimate of the number of birds present can be obtained; a smaller SD greatly improves the ability to detect changes in numbers between counts.
While the coefficient of variation, obtained by Rappoldt et al., may appear surprisingly high, it is consistent with others determined in the laboratory (28%: Erwin, 1982) and in the field with shorebirds (30–80%, depending on site: Rogers et al., 2006).
All count data were entered into pre-prepared Excel spreadsheets containing separate worksheets for each survey area (which included the sub-areas and their survey sites) and a Summary worksheet. Disturbance data were also collated into an Excel spreadsheet for analysis.
Our analysis mainly focused on survey data collected during the first winter to illustrate the important types of information that can be obtained from systematic surveys. However, the practical difficulties encountered during the 2009/2010 winter in achieving the preferred 2–3-day surveys provided an important opportunity to assess the ability of the methodology to provide reliable results when surveys are conducted over extended periods. This assessment is included in the Discussion.
Results
Water levels
The daily Shengjin Lake water levels at the Huangpen Sluice during the the 2008/2009 waterbird wintering period are shown in Fig. 3; data on rainfall and timing of surveys are also included.
Figure
3.
Water levels (Wusong datum) for Shengjin Lake (continuous line) at the Huangpen Sluice gauge from 1 October 2008 to 16 April 2009; the seven survey periods (mid-dates; open triangles) and rainfall data from 23 October to 15 April 2009 are included (adapted from Fig. 5 in Cheng et al., 2009).
Lake water levels fell after the flood season to a mid-winter low of 8–9 m. A large discharge from the Three Gorges Dam in early November caused the lake water level to rise quickly to more than 12 m, before falling to more normal levels by early December. High catchment rainfall in mid- to late February led to a rise in the lake level to 11 m, following which it fell steadily during March.
Analysis of the 2008/2009 winter waterbird counts
The results of the seven counts are listed in Table 1, with species sorted according to their maximum counts over the seven surveys. Maximum total numbers occurred when water levels were at their lowest (8–9.5 m).
Table
1.
Species counts for the seven surveys conducted in the winter of 2008/2009, sorted according to the maximum species counts during the winter (adapted from Table 1 in Cheng et al., 2009)
The count data have been used to determine the key waterbird groups and species present at the lake, to identify important waterbird areas and to compare the early-February 2009 and mid-February 2010 counts with similar counts made in February 2004 and 2005.
Adjustments were made to some species totals in Surveys 3 and 4 to allow for birds missed due to poor visibility; the corrections were based on counts made during the 1–2 days following the surveys and included increasing the number of some species and the inclusion of counts for completely missed species. The systematic survey methodology developed for the surveys facilitated these adjustments because of the use of sub-areas and the detailed knowledge of environmental conditions that applied during the surveys.
Key waterbird groups and species
The waterbird community at Shengjin Lake is dominated by Anatidae, which represented 86% of all waterbirds based on maximum numbers counted, followed by shorebirds (6.1%), gulls and terns (2.7%) and herons and egrets (1.9%).
A total of 13 key species were identified during the 2008/2009 winter period using the criteria that they were 1) present in numbers greater than 1% of their flyway population (Wetlands International, 2006) or 2) globally threatened (IUCN, 2009). These species are Black Stork (Ciconia nigra), Oriental Stork (Ciconia boyciana), Eurasian Spoonbill (Platalea leucorodia), Tundra Swan (Cygnus columbianus), Swan Goose (Anser cygnoides), Bean Goose (Anser fabalis), Greater White-fronted Goose (Anser albifrons), Lesser White-fronted Goose (Anser erythropus), Falcated Duck (Anas falcata), Baikal Teal (Anas formosa), Siberian Crane (Grus leucogeranus), Hooded Crane (Grus monachal) and Spotted Redshank (Tringa erythropus).
Shengjin Lake held the largest recorded concentration of Bean Geese in the Yangtze River floodplain (60% of the global serrirostris subspecies population) in mid-December 2008.
Key waterbird areas
The numbers of waterbirds counted in each survey area during the seven surveys are shown in Table 2. Survey area E held considerably more birds than any other area in all surveys except for the final count in mid-April, when very few waterbirds remained, supporting 40–72% of all waterbirds in the four highest counts. The numbers in survey area A, the second most important region, varied little during the peak period, while those in survey areas C and D were at a maximum in early-February when water levels were at their winter low.
Table
2.
Number of waterbirds counted in each survey area during the seven surveys in the winter of 2008/2009 (adapted from Table 5 in Cheng et al., 2009)
Analysis of the waterbird distribution data enabled 18 key waterbird areas to be identified (Fig. 4). These areas were selected on the basis that they held important concentrations of waterbirds at some stage during the winter, either in terms of total numbers, a high number of an individual species or the presence of globally threatened species (Box 2).
Figure
4.
Key waterbird areas identified during the 2008/2009 winter (adapted from Fig. 8 in Cheng et al., 2009)
Comparison of 2008/2009 and 2009/2010 counts with 2003/2004 and 2004/2005 counts
Comprehensive counts of Shengjin Lake were conducted in February 2004 and February 2005 (Barter et al., 2004, 2006). These provided a useful baseline against which to compare the equivalent February counts in the 2008/2009 and 2009/2010 winters. Water levels were reasonably similar for the four counts, ranging from 8.1–9.7 m, approximately the range which supported the highest number of waterbirds during the 2008/2009 and 2009/2010 winters.
Comparisons between the four counts (Table 3) show that there are two key species in which numbers have decreased greatly between the two periods: Tundra Swan and Swan Goose, and two whose abundance has increased considerably: Bean Goose and Greater White-fronted Goose.
Table
3.
Comparison of counts of four species and total counts in 2004, 2005, 2009 and 2010 (adapted from Table 4 in Cheng et al., 2009)
The magnitude and location of some of the major potential disturbance pressures occurring in the winter of 2009/2010 are shown in Fig. 5 (Note: second winter data used because of availability of more extensive data sets). The total number of people is not large, peaking at ~90 in early December; the highest numbers occurred in the Upper Lake in the same regions as major buffalo concentrations. The highest number of buffalos were found in the Upper Lake in the same areas as major geese concentrations, which is to be expected for they are foraging on the same food resource (Carex sp.). Most boat movements occurred in early January, associated with the peak fish-harvesting period; major boat activity was confined to the northern and western shores of the Upper Lake which are free of nets, thus allowing boat fishing to take place. Large domestic geese and duck flocks were present at a number of locations, generally close to major waterbird concentrations where most food was available; numbers reached low levels in the lead up to the Spring Festival period following slaughter for human consumption. All these threats are mainly concentrated in the Upper Lake which is the most important region for waterbirds in Shengjin Lake (Table 2).
Figure
5.
Graphs show maximum numbers of (a) people, (b) buffalos, (c) moving boats, and (d) domestic geese and ducks encountered during surveys conducted in the 2009/2010 winter; for clarity, maps only show distributions of highest numbers encountered during the winter period (i.e. > 10 people, ≥10 buffalos, ≥5 moving boats and ≥100 domestic geese and ducks).
The overall importance of Shengjin Lake is demonstrated by the occurrence of high numbers of waterbirds and the presence of 13 key species (either globally threatened or present in numbers of international importance). The highest number of waterbirds occurs when water levels are low. A total of 18 key waterbird areas were identified within the lake. Comparison with 2004 and 2005 counts shows that numbers of two key species have declined considerably and two have increased significantly. Disturbance data indicate that the highest number of waterbirds occur in the most threatened areas.
Discussion
The significant benefits that arise from using a systematic method to survey waterbirds and their habitats are clearly demonstrated by the quality of the data obtained on i) species numbers and how these change during the winter; ii) community structures; iii) key species; iv) important habitats and v) threatening processes. The availability of environmental and disturbance data greatly assisted with data adjustment and interpretation.
We believe that the counts at Shengjin Lake have a high level of accuracy and precision because i) surveying mostly took place over open expanses of water and marshes where flocking waterbirds are generally highly visible and readily counted using high quality optical equipment; ii) virtually all surveys took place from elevated positions, e.g. dykes, tall buildings and hills (particularly in the Upper Lake) and iii) observers were experienced, having taken part in many surveys and research activities at the lake. We believe it will be possible to use the data in the future, when a larger longitudinal set is available, to test whether differences exist between inter- and intra-seasonal counts, thus providing a good basis for the development of a statistically robust monitoring program at Shengjin Lake.
Box 2 Bases for the selection of the 18 key areas
Survey area A
1. Supports large numbers of Bean Geese; also important for Oriental Stork, Swan Geese, herons, egrets and spoonbills at lower water levels.
2. Important foraging area for Bean Geese when water levels are high.
3. Large numbers of Bean Geese and Greater White-fronted Geese feed on Carex meadows (second most important goose region in Shengjin Lake); important for Tundra Swans, Oriental Storks, Eurasian Spoonbills, ducks and shorebirds when water levels fall to provide shallow water.
Survey area B
4. Supports large numbers of Swan Geese, Bean Geese and Tundra Swans when water levels fall, allowing foraging on Vallisneria tubers and Carex.
5. Important foraging area for Tundra Swans and Bean Geese, feeding mainly on underwater vegetation.
Survey area C
6. Important for Tundra Swans (continuation of key site 5).
7–9. Bays are very important for Swan Geese, Bean Geese and Tundra Swans feeding mainly on underwater vegetation, especially when lake water levels are high and before the bays are drained for fish harvesting.
10. Important for Swan Geese, Bean Geese and Tundra Swans when lake water levels are high, feeding over exposed mud flats inside dam.
11. Important for Bean Geese and Tundra Swans when water levels are low, feeding mainly on underwater vegetation.
12. Important for Bean Geese and Tundra Swans, feeding mainly on underwater vegetation.
Survey area D
13. Important roosting area for ducks, especially Falcated Duck and Baikal Teal.
Survey area E
14. Important for Bean Geese and ducks, feeding mainly on underwater vegetation.
15. Rice paddies are most important foraging area for Hooded Cranes feeding mainly on spilt rice and winter wheat; also used by Bean Geese.
16. Rice paddies are important foraging areas for Hooded Cranes feeding mainly on spilt rice and winter wheat; also used by Bean Geese.
17. Most important waterbird region at Shengjin Lake. Carex meadows support highest numbers of Bean Geese and Greater White-fronted Geese in Shengjin Lake; also large numbers of Tundra Swans, ducks and shorebirds feed in the lake and on the mud flats at low water levels.
18. Rice paddies support large numbers of foraging Hooded Cranes feeding mainly on spilt rice and winter wheat; also used by Bean Geese.
Given the advantageous circumstances under which the winter 2008/2009 surveys were generally conducted, the main factor which might have affected count accuracy was most likely bias due to rates of detection of less than 100%, particularly of cryptic and less common species; this will lead to underestimation of numbers. Thus, it is important to obtain estimates of rates of detection under different conditions in order that count accuracy be further improved; Thompson (2002) discussed a number of approaches to measure rates of detection, including double sampling and the double-observer approach. These techniques need investigation to determine whether they can provide satisfactory rates of detection under the conditions at Shengjin Lake. It is also important to determine whether species identification errors are significant; Hull et al. (2010) found that up to 23% of similar raptor species were misidentified during censuses and there seems to be a comparable potential for misidentification of waterbirds, particularly of some duck and shorebird species.
The importance of conducting regular surveys throughout the winter is shown by variations in total numbers and number of species from survey-to-survey. Frequent surveys throughout the winter also provide useful information on the timing of migration arrivals and departures, as well as of influxes. For this reason, surveys in the 2009/2010 winter were scheduled for approximately every two weeks starting in mid-October. A crude analysis of the 2009/2010 count data (Meng, 2010) showed that community structure, peak numbers and movements were similar to those obtained in the previous winter, e.g. 17 out of the 20 most common species in 2008/2009 were in the top 20 for 2009/2010; peak total numbers occurred in December in both winters and were similar in magnitude (i.e. 71423 in 2008/2009; 63639 in 2009/2010); the maxima for 13 of the 20 species occurred within the same 4-week period in both winters and were similar in magnitude for 10 species (i.e. 2009/2010 numbers were within ±25% of those in 2008/2009). These initial results indicate that the waterbird community is relatively stable from year-to-year. It also seems that the methodology is capable of providing consistent results from year-to-year, despite widely varying survey conditions; however, additional data are required to confirm this.
Collection of environmental information is very important for data interpretation and assessment of which factors could be driving changes in species abundance. For example, water levels are probably the most important variable determining absolute numbers; the greatest number of birds were present in both winters when water levels were in the 8–9.5 m range, presumably because of the increased areas of shallow water and grazing marshes. Knowledge of water levels is also very important when comparing surveys across seasons and years; without this information, incorrect conclusions concerning causes of changes in abundance may be drawn. Other environmental information on weather, viewing distances and count completeness will assist the analyst in understanding and explaining unexpected variations in count numbers.
Similarly, disturbance information can help in understanding the nature, distribution and magnitude of threatening processes. For example, the impact of the extensive netted fishponds in the western and southern parts of the Lower Lake can be seen in the relative lack of key areas in this region (Fig. 4, survey areas B and C), those which did occur were mainly in the fringing bays. In contrast, the large un-netted open area (13) in survey area D supported large flocks of roosting ducks. Knowledge of threatening processes is important for management purposes, e.g. controlling human access to key waterbird areas and identifying important research directions, e.g. investigating buffalo-goose interaction on grazing meadows. The disturbance data collected to date will also provide a baseline against which to measure changes in future disturbance levels, allowing an assessment to be made of their impacts on the number of waterbirds and their distributions.
It is very important for conservation purposes to determine the reasons for changes in abundance and distribution which have been highlighted by comparative data for key species over the 2004/2005–2009/2010 period. Investigations have already commenced into potential explanations of these changes, e.g. Swan Geese (Zhang et al., 2010), Bean Geese (Zhao et al., 2010), Greater White-fronted Geese (MJ Zhao, unpublished data) and Tundra Swans (PH Cong, unpublished data). The changing abundances and distributions of key species is an important indicator of wetland health (Fox et al., 2010).
Given the growing threats to Yangtze River floodplain wetland habitats and the consequent decline in abundance and changing distributions of key species, it is critically important to develop a floodplain-wide waterbird population monitoring program. This will not be a simple task for it requires the establishment of similar waterbird survey programs at a large number of wetlands. It is unlikely that the resources to achieve this are currently available (e.g. sufficient skilled surveyors to develop survey programs and conduct frequent surveys throughout the winter). However, unless such a methodology is adopted and rigorously implemented it will be impossible to determine whether statistically significant changes are taking place in numbers and distributions of waterbirds across the floodplain.
It will be highly desirable to estimate the power of any proposed monitoring program to detect temporal population trends across multiple sites (Haslem et al., 2008). This can be done with software such as MONITOR (www.mbr-pwrc.usgs.gov/software/monitor.html) which uses inputs such as proposed number of survey sites, mean species abundance, variability at each site and the number of surveys (annual and seasonal), to estimate the ability of the program to detect predetermined trends (e.g. decline of 5% per year) at a given significance level (e.g. 0.05%). By using existing data on number of species and variability (e.g. Barter et al., 2004, 2006), it should be possible to determine the power of alternative programs to detect changes at the required level of significance.
Acknowledgements
Thanks to Songtao Dou, Xiuli Yang and Jing Liu and the Anhui Shengjin Lake National Nature Reserve staff and local fishermen for assisting with the fieldwork and/or helping with the preparation of the manuscript. The study was supported by the National Natural Science Foundation of China (Grant Nos. 31071941 and 30940010), the Chinese Academy of Sciences Innovation Project (Grant No. KZCX2-YW-QN503) and the University of Science and Technology of China Graduate School Educational Innovation Base.
Appendix 1.
Directions for survey area A following GPS track
1) Turnoff main road at (T1), to E of sluice, and drive S via turnoffs at (T2) and (T3) and park car about 200 m down narrow gravel road to dam.
2) Walk to survey site 1 and survey region inside dam (sub-area a1), especially the peninsular region; also survey bays in far S of sub-area a3.
3) Return to survey site 2 and survey reservoir to E (sub-area a2).
4) Return to car and drive via turnoffs (T3) and (T2) to Yanwo Management Station (survey site 3). From roof, survey regions to NW, W, SW and S (sub-area a3).
5) Drive via turnoffs (T2), (T1), (T4) and (T5) and park car at (T6). Survey fishpond to S (sub-area a4) from survey site 4. Then walk to survey site 5 and survey region to N, W and SW (part sub-area a6).
6) Walk along small dyke to survey site 6 and survey fish pond to S (sub-area a5).
7) Return to car and drive via turnoff (T5) to survey site 7 and survey region to SE, S and SW (part sub-area a6). Note that survey sites along dyke are flexible depending on location of birds and viewing conditions.
8) Continue along dyke to survey site 8 and survey region to SE, S and SW (part sub-area a6).
9) Continue along dyke to survey site 9 and survey region to NE, E and SE (part sub-area a6).
10) Continue along dyke to survey site 10 and survey region to E, S and SE (sub-area a7).
11) Continue along dyke to survey site 11 and survey region to E (sub-area a8).
Appendix 2.
Site survey form
Name:
Wetland:
Survey area/Survey site No.:
Contact details (Phone/email):
Date:
Start:
Finish:
Survey type:
Viewing conditions:
Wind direction:
Wind speed:
Water level:
Temperature:
Rain?
Cloud cover:
Complete?
Accurate?
Disrupted?
Total number of observers:
Distance to most of birds:
Max. distance of birds:
Years of experience:
Total waterbirds:
Total species:
Species
Component counts
Total
Comment
Little Grebe
Great-crested Grebe
Great Cormorant
Grey Heron
Eastern Great Egret
Little Egret
Eurasian Bittern
Black Stork
Oriental Stork
Eurasian Spoonbill
Tundra Swan
Swan Goose
Bean Goose
Greater White-fronted Goose
Lesser White-fronted Goose
Greylag Goose
Bar-headed Goose
Ruddy Shelduck
Common Shelduck
Eurasian Wigeon
Falcated Duck
Gadwall
Common Teal
Northern Mallard
Spot-billed Duck
Northern Pintail
Northern Shoveler
Common Pochard
Tufted Duck
Smew
Goosander
Siberian Crane
White-naped Crane
Common Crane
Hooded Crane
Brown Crake
White-breasted Waterhen
Common Moorhen
Common Coot
Pied Avocet
Northern Lapwing
Grey-headed Lapwing
Grey Plover
Long-billed Plover
Kentish Plover
Common Snipe
Black-tailed Godwit
Eurasian Curlew
Spotted Redshank
Common Greenshank
Green Sandpiper
Common Sandpiper
Dunlin
Herring Gull
Common Black-headed Gull
UID egrets
UID spoonbills
UID geese
UID ducks
UID cranes
UID rails
UID shorebirds
UID snipe
UID gulls
UID waterbirds
Number of the following observed from the survey position:
People moving (not in boats):
Goats:
Motorbikes and bicycles:
Buffaloes (count all seen):
Boats at anchor:
Chicken flocks:
Dogs (in or at edge of wetland):
Boats moving:
Domestic duck flocks:
Other:
Cars/Trucks:
Domestic geese flocks:
Number of disturbance flights during survey period:
If the total number of waterbirds in this area was notably different than usual, please provide any possible explanation here:
Have there been noticeable changes in the habitat since the last visit? (please describe)
Comments:
Appendix 3.
Instructions for completing site survey form
Name:
First and last name of survey leader
Wetland:
The name of the wetland that the survey area is a part of, e.g. Shengjin Lake
Survey Area/Site No.:
Name of survey area and survey site number, e.g. A-27
Contact details:
Phone number and e-mail address
Date:
Day/month/year (dd/mm/yyyy)
Start Time:
Use 24 hour clock to nearest 15 minutes (i.e. 14:00 = 2:00 pm)
Finish Time:
Use 24 hour clock to nearest 15 minutes
Survey type:
L = land, B = Boat
Viewing conditions:
Enter code on scale of (0 to 5): where 0 = Excellent conditions with good light, can clearly see individual feathers on close birds, to a 5 = Extremely poor conditions where you can barely see the birds due to glare from the sun, too dark, extreme wind etc.
Wind direction:
Enter compass direction from which the wind is coming, i.e., N, NE, SE, etc. Variable = VAR
Wind Speed:
Enter code (0 to 6): 0 = 0 to 5 km·h–1 (wind not felt on face), 1 = 6–11 kph (wind felt on face), 2 = 12–20 kph (leaves, small twigs in constant motion, light flag extended), 3 = 21–29 km·h–1 (raises dust, small branches in motion), 4 = 30–39 km·h–1 (small trees in leaf sway), 5 = 40–49 km·h–1 (large branches in motion, whistling heard in wires), 6 = greater than 50 km·h–1 (resistance felt while walking)
Water level:
Use depth on Shengin Lake guage near Huangpen Sluice.
Temperature:
Obtain from climate meter records at Field Centre.
Rain?:
Enter code (0 to 3): 0 = not raining, 1 = light rain, 2 = moderate rain, 3 = heavy rain
Cloud cover:
Enter code (0 to 4): 0 = no cloud, 1 = 25% cloud, 2 = 50% cloud, 3 = 75% cloud, 4 = 100% cloud
Complete?
Was the entire area covered? YES or NO (if NO please note on map the area covered)
Accurate?:
Was the count accurate? YES or NO (if NO please add comment)
Disrupted?:
Did anything disrupt the survey? (if YES please add comment)
Total number of observers:
Record the number of observers taking part in the count.
Distance to most birds:
Distance in metres to largest concentration of birds
Max. Distance of birds:
Distance in metres to furthest birds counted.
Years of experience
Number of years that most experienced counter has been counting waterbirds.
Total waterbirds:
Total number of waterbirds recorded from survey site
Total waterbird species:
Number of different waterbirds species counted from survey site.
Component counts It is important to record the number of each species using component counts. So during a count you might record a total of 1000 Common Teals, but unless they were all together or you counted them all at once with one estimate, they should be recorded as 400 / 9 / 3 / 27 / 60 / 230 / 1 / 270. Recording data this way allows us to reduce the variance of the counts for individual species. Put an F after any count where the birds were flying through the count area, and were not seen using any of the count area during the visit. For example, your data might look like this: Common Teal 11 / 35 / 444 / 325 / 77 / 10 Northern Mallard 40 / 12000 / 780 F / 848 / 7 / 39 Dunlin 57 / 5 / 2 / 5000 / 99 / 878 Mark the comment box for each species as needed.
Number of the following observed from the survey position:
People moving (not in boats):
Goats:
Motorbikes and bicycles:
Buffaloes (count all seen):
Boats at anchor:
Chicken flocks:
Dogs (in or at edge of wetland):
Boats moving:
Domestic duck flocks:
Other:
Cars/Trucks:
Domestic geese flocks:
Number of disturbance flights during survey period:
Number of times waterbirds flew in apparent response to disturbance during the survey (including from counters).
If the total number of waterbirds in sub-area was notably different than usual, please provide any possible explanation here:
(Often this is the kind of data that cannot be guessed at during data analysis, but can point further study in the right direction if there is some indication of what is thought to be driving changes in populations)
Have there been noticeable changes in the habitat since the last visit? (please describe)
Comments: Add anything of interest, especially anything that may have affected the survey. Do you suspect that there were large numbers of birds that were hidden from view. Did conditions affect the survey? Any thoughts on how things could be improved (are boats needed etc)?
An B, Zhang LX, Liu NF, Browne S, Ruan LZ, Song S. 2009. Phylogeography of Tibetan snowcock (Tetraogallus tibetanus) in Qinghai-Tibetan Plaetau. Mol Phylogenet Evol, 50: 526–533
Avise JC. 1994. Molecular Markers, Natural History and Evolution. Chapman & Hall, New York
Avise JC, Walker D. 1998. Pleistocene phylogeographic effects on avian populations and the speciation process. Proc R Soc Ser B-Biol, 265: 457–463
Belkhir K, Borsa P, Chikhi L, Raufaste N, Catch F. 2004. GENETIX (version 4.02), Software under WindowsTM for the genetics of the populations Laboratory Genome, Populations, Interactions, CNRS UMR 5000, University of Montpellier Ⅱ, Montpellier, France
Chen Q, Chang C, Liu NF. 1999. Mitochondrial DNA introgression between two parapatric species of Alectoris. Acta Zool Sin, 45(4): 456–463 (in Chinese)
Chen YK, Zhao ZL, Liu NF. 2006. Genetic structure of Przewalski's Rock Partridge (Alectoris magna) population in the Longdong Loess Plateau, China. Biochem Genet, 44: 209–221
Chistiakov DA, Hellemans B, Volckaert FAM. 2006. Microsatellites and their genomic distribution, evolution, function and applications: a review with special reference to fish genetics. Aquaculture, 255: 1–29
Comes HP, Kadereit JW. 1998. The effect of Quaternary climatic changes on plant distribution and evolution. Trends Plant Sci, 3: 432–438
Di Rienzo A, Peterson AC, Garza JC, Valdes AM, Slatkin M, Freimer NB. 1994. Mutational processes of simple-sequence repeat loci in human populations. Proc Natl Acad Sci USA, 91: 3166–3170
El Mousadik A, Petit RJ. 1996. High level of genetic differentiation for allelic richness among populations of the argan tree[Argania spinosa (L. ) Skeels] endemic to Morocco. Theor Appl Genet, 92: 832–839
Excoffier L, Laval G, Schneider S. 2005. ARLEQUIN, version3.0: an integrated software package for population genetics data analysis. Evol Bioinfor Online, 1: 47–50
Frankel OH, Soule ME. 1981. Conservation and Evolution. Cambridge University Press, Cambridge, England
Gaggiotti OE, Excoffier L. 2000. A simple method of removing the effect of a bottleneck and unequal population sizes on pairwise genetic differences. Proc R Soc Lond B, 267: 81–87
Gerloff U, Schlötterer C, Rassmann K, Rambold I, Hohmann G, Fruth B, Tautz D. 1995. Amplification of hypervariable simple sequence repeats (microsatellites) from excremental DNA of wild living bonobos (Pan paniscus). Mol Ecol, 4: 515–518
Goudet J. 2001. FSTAT. A program to estimate and test gene diversities and fixation indices, (Version 2.9.3). Updated from Goudet (1995)
Hamrick JL, Godt MJW. 1989. Allozyme diversity in plant species. In: Brown AHD, Clegg MT, Kahler AL, Weir BS (eds) Plant Population Genetics, Breeding, and Genetic Resources. Sinauer, Sunderland, MA, pp 43–63
Hamrick JL, Godt MJW, Sherman-Broyles SL. 1992. Factors influencing levels of genetic diversity in woody plant species. New Forest, 6: 95–124
Hartl DL, Clark AG. 1997. Principles of Population Genetics. Sinauer, Sunderland, MA
Heuertz M, Hausman JF, Hardy OJ, Vendramin GG, Frascaria-Lacoste N, Vekemans X. 2004. Nuclear microsatellites reveal contrasting patterns of genetic structure between western and southeastern European populations of the common ash (Fraxinus excelsior L. ). Evolution, 58: 976–988
Hewitt GM. 2000. The genetic legacy of the Quaternary ice ages. Nature, 405: 907–913
Hewitt GM. 2004. Genetic consequences of climatic oscillations in the Quaternary. Philos T R Soc Lon B, 359: 183–195
Hou P, Wei M, Zhang LX, Liu NF. 2002. Genetic structure of edge population in Przewalski's rock partridge (Alectoris magna). Acta Zool Sin, 48(3): 333–338 (in Chinese)
Huang ZH, Liu NF. 2007. Phylogeography of rusty-necklaced partridge (Alectoris magna) in northwestern China. Mol Phylogenet Evol, 43: 379–385
Johnsgard PA. 1988. The Quails, Partridges, and Francolins of the World. Oxford University Press, Oxford
Liu NF, Huang ZH, Wen LY. 2004. Subspecies divergence of Przewalski's rock partridge (Alectoris magna) description of a new subspecies. Acta Zootaxon Sin, 29(3): 600–605 (in Chinese)
Liu NF, Wang XT, Luo WY, Chang C. 1988. Avifauna study of the Liupan Mountain in Ningxia. J Lanzhou Univ Nat Sci Ed, 24: 63–75 (in Chinese)
Luikart G, Cornuet JM. 1997. Empirical evaluation of a test for identifying recently bottlenecked populations from allele frequency data. Conserv Biol, 12: 228–237
Madge S, McGowan P. 2002. Pheasants, Partridges and Grouse: A Guide to the Pheasants, Partridges, Quails, Grouse, Guineafowl, Buttonquails and Sandgrouse of the World. Christopher Helm, London
Mantel N. 1967. The detection of disease clustering and a generalized regression approach. Cancer Res, 27: 209–220
Maruyama T, Fuerst PA. 1985. Population bottlenecks and nonequilibrium models in population genetics. Ⅱ. Number of alleles in a small population that was formed by a recent bottleneck. Genetics, 111: 675–689
Michalakis Y, Excoffier L. 1996. A genetic estimation of population subdivision using distance between alleles with special reference to microsatellite loci. Genetics, 142: 1061–1064
Nei M. 1972. Genetic distance between populations. Am Nat, 106: 283–292
Nei M. 1987. Molecular Evolutionary Genetics. Columbia University Press, New York
Nei M, Maruyama T, Chakraborty R. 1975. The bottleneck effect and genetic variability in populations. Evolution, 29: 1–10
Nei M. 1978. Estimation of average heterozygosity and genetic distance from a small number of individuals. Genetics, 89: 583–590
Prentice HC, Lonn M, Lefkovitch LP, Runyeon H. 1995. Associations between alleles frequencies in Festuca ovina and habitat variation in the alvar grasslands on the Baltic island of Oland. J Ecol, 83: 391–402
Pritchard JK, Stephens M, Donnelly P. 2000. Inference of population structure using multilocus genotype data. Genetics. 155: 945–959
Ramamoorthi J, Thilagam K, Sivaselvam SN, Karthickeyan SMK. 2009. Genetic characterization of Barbari goats using microsatellite markers. J Vet Sci, 10(1): 73–76
Raymond M, Rousset F. 1995a. Genepop version 12: population genetics software for exact tests and ecumenicism. J Hered, 86: 248–249
Raymond M, Rousset F. 1995b. An exact test for population differentiation. Evolution, 49: 1280–1283
Rousset F. 1997. Genetic differentiation and estimation of gene flow from Fstatistics under isolation by distance. Genetics, 145: 1219–1228
Slatkin M. 1987. Gene flow and the geographical structure of natural populations. Science, 236: 787–792
Slatkin M. 1993. Isolation by distance in equilibrium and non-equilibrium populations. Evolution, 47: 264–279
Slatkin M. 1995. A measure of population subdivision based on microsatellite allele frequencies. Genetics, 139: 457–462
Sun HY, Cao ZW, Jiang HL, Yu LG. 1998. Brooding and growth of the younger Perdix dauuricae. Forest Sci Technol, 23(4): 28–30 (in Chinese)
Sun HY, Ge DN, Hong YT, Cao ZW. 1996. Research about domesticating and breeding technology of Daurian Partridge (Perdix dauuricae). Forest Sci Technol, 21(4): 32–35 (in Chinese)
Takezaki N, Nei M. 1996. Genetic distances and reconstruction of phylogenetic trees from microsatellite DNA. Genetics, 144: 389–399
Tautz D. 1989. Hypervariability of simple sequences as a general source for polymorphic DNA markers. Nucleic Acids Res, 17: 6463–6471
Turner PC, McLennan AG, Bates AD, White MRH. 2003. Instant Notes in Molecular Biology, 2nd edn. Science Press, Beijing
Weir BS, Cockerham CC. 1984. Estimating F-statistics for the analysis of population structure. Evolution, 38: 1358–1370
Woodruff DS. 1989. The problems of conserving genes and species. In: Weston D, Pearl M (eds) Conservation for the Twenty First Century. Oxford University Press, New York, pp 76–88
Wright S. 1943. Isolation by distance. Genetics, 28: 114–138
Zhang ZW, Liang W, Sheng G. 1994. Studies on the nest site selection of Daurian partridge. Zool Res, 15(4): 37–43 (in Chinese)
Zhang ZW, Wu YC. 1992. The Daurian Partridge (Perdix dauuricae) in north-central China. Gibier Faune Sauvage, 9: 591–595
Zhang ZW, Liang W. 1997. Breeding ecology of Daurian Partridge (Perdix dauuricae) in Shanxi. Chinese J Zool, 2(2): 23–25 (in Chinese)
Zhao ZJ, Zhang S, Feng KF. 1992. The biology of the Daurian Partridge (Perdix dauuricae suschkini) in northeastern China. Gibier Faune Sauvage, 9: 597–604
Table
1.
Species counts for the seven surveys conducted in the winter of 2008/2009, sorted according to the maximum species counts during the winter (adapted from Table 1 in Cheng et al., 2009)
Table
2.
Number of waterbirds counted in each survey area during the seven surveys in the winter of 2008/2009 (adapted from Table 5 in Cheng et al., 2009)


 DownLoad:
DownLoad:









 Email Alerts
Email Alerts RSS Feeds
RSS Feeds