Yufei Dai, Qingxian Lin, Wenzhen Fang, Xiaoping Zhou, Xiaolin Chen. 2015: Noninvasive and nondestructive sampling for avian microsatellite genotyping: a case study on the vulnerable Chinese Egret (Egretta eulophotes). Avian Research, 6(1): 24. DOI: 10.1186/s40657-015-0034-x
Citation:
Yufei Dai, Qingxian Lin, Wenzhen Fang, Xiaoping Zhou, Xiaolin Chen. 2015: Noninvasive and nondestructive sampling for avian microsatellite genotyping: a case study on the vulnerable Chinese Egret (Egretta eulophotes). Avian Research, 6(1): 24. DOI: 10.1186/s40657-015-0034-x
Yufei Dai, Qingxian Lin, Wenzhen Fang, Xiaoping Zhou, Xiaolin Chen. 2015: Noninvasive and nondestructive sampling for avian microsatellite genotyping: a case study on the vulnerable Chinese Egret (Egretta eulophotes). Avian Research, 6(1): 24. DOI: 10.1186/s40657-015-0034-x
Citation:
Yufei Dai, Qingxian Lin, Wenzhen Fang, Xiaoping Zhou, Xiaolin Chen. 2015: Noninvasive and nondestructive sampling for avian microsatellite genotyping: a case study on the vulnerable Chinese Egret (Egretta eulophotes). Avian Research, 6(1): 24. DOI: 10.1186/s40657-015-0034-x
Key Laboratory of the Ministry of Education for Coast and Wetland Ecosystems, School of Life Sciences, College of the Environment and Ecology, Xiamen University, Xiamen 361102, China
Noninvasive and nondestructive DNA sampling techniques are becoming more important in genetic studies because they can provide genetic material from wild animals with less or even without disturbance, which is particularly useful for the study of endangered species, i.e., birds. However, nondestructively and noninvasively sampled DNA may, in some cases, be inadequate in the amount and quality of the material collected, which can lead to low amplification success rates and high genotyping errors.
Methods
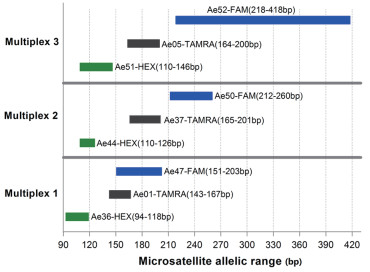
In this study, noninvasive (eggshell swab, shed feather and feces), nondestructive (plucked feather and buccal swab) and invasive (blood) DNA samples were collected from the vulnerable Chinese Egret (Egretta eulophotes). DNA concentrations, PCR amplification success and microsatellite genotyping errors of different sample types were evaluated and compared to determine whether noninvasive and nondestructive samples performed as well as invasive samples in our experimental procedures.
Results
A total of 159 samples were collected in the field. Among the different sample types, the highest DNA concentrations (154.0–385.5 ng/μL) were obtained from blood. Those extracted from fecal samples were the lowest, ranging from 1.25 to 27.5 ng/μL. Almost all of the DNA samples, i.e., 95.59 %, were successfully amplified for mtDNA (n = 152) and 92.76 % of mtDNA samples were successfully genotyped for at least five of the nine microsatellite loci tested (n = 141). Blood samples and buccal swabs produced reliable genotypes with no genotyping errors, but in feces, allelic dropouts and false alleles occurred in all nine loci, with error rates ranging from 6.67 to 38.10 % for the dropouts and from 6.06 to 15.15 % for the false alleles.
Conclusions
These results indicate that both nondestructive and noninvasive samplings are suitable for avian microsatellite genotyping, save for fecal DNA. However, we should remain cautious of the appearance of genotyping errors, especially when using noninvasive material.
Breeding, migration, and moulting are considered the three-main energy-demanding events in the yearly cycle of birds (Newton 2008) and usually studied separately due to their isolated occurrence (Newton 2009). The importance of the moult in the life cycle of birds of prey for advancing our understanding of the ecology of each species has been recently highlighted by Zuberogoitia et al. (2018). In several migrating species moulting leads to age-dependent migration strategies (Cristol et al. 1999; Zenzal and Moore 2016) and these differences are especially pronounced among long-distance migrants. Among passerines, first-year birds migrate earlier than adults. The opposite occurs in species in which adults suspend or postpone moult until they reach their wintering grounds (Schifferli 1965; Payevski 1994; Woodrey and Chandler 1997; Woodrey and Moore 1997; Jakubas and Wojczulanis-Jakubas 2010). Long distance migrant raptors typically avoid moulting during migration (Berthold 1975; Newton 1979) such as in the case of Osprey (Pandion haliaaetus; Ferguson-Lees and Christie 2001). In small raptors the time required to grow a new feather is 2-3 weeks while it is much longer in large raptors such as vulture species (Zuberogoitia et al. 2018). In the case of damaged feathers, raptors can replace them much faster than in the usual time of moulting (Ellis et al. 2016). However, there are species that do not stop moulting during long-distance movements (Kjellén 1992). For example, it has been found that Palearctic Circus sp. moult during migration and eventually slow the moulting process (Arroyo and King 1996). Moreover, we can find the "suspended moult" strategy which is defined as a moult interrupted temporarily and subsequently resumed (Baker 2016). This strategy frequently involves individuals in late breeding attempts having to interrupt their normal moult cycle in order to migrate.
In Western Marsh Harriers (WMHs), females started moulting during incubation (May-June) while males start up to 6 weeks later (Forsman 1999). During chicks rearing period WMH may suspend moult for better hunting efficiency while they end moulting in late October-early November (Forsman 1999). During spring migration Gorney and Yom (2001) showed that adult WMHs perform their journey with non-active moult in their flight feathers.
Unfortunately, moulting patterns in relation to migration strategies are poorly investigated in raptor species despite it is well known that long-distance migrant Accipitriformes need a large amount of energy to overcome barriers during migration where they must use flapping powered flight (Agostini et al. 2015). The aim of this work is to confirm and to describe the active moult of WMH's flight feathers during the spring migration across the Strait of Messina.
Methods
Fieldwork
From August 13th to September 30th 2016 we collected data on moulting Western Marsh Harriers migrating along the Mid-Mediterranean Flyway (Fig. 1). Fieldwork was carried out on the continental side of the Strait of Messina, a hotspot for soaring birds (Panuccio et al. 2005; Agostini et al. 2017; Pastorino et al. 2017). We used two watch points both located on the Aspromonte Plains, a flat highland a few kilometers inland of the Strait of Messina, on the western side of the Apennines ridge. The highland lies between 1000 and 1200 m a.s.l. with a landscape altered by human agricultural activity. One of the watch points was located in the middle of the highland while the other one was located close the mountain slopes. Observations were carried out daily from sunrise to sunset.
Figure
1.
Geographical context of the study area, the Strait of Messina (SM). The asterisk indicates the Aspromonte plain
We used high-quality photos taken in the field from which moult patterns can be inferred (Snyder et al. 1987; Zuberogoitia et al. 2016; Vieira et al. 2017). Digital cameras (i.e. Panasonic Lumix DMC-FZ72, FZ300) were equipped with lens with 8-60 × magnification. Photography data collection was supported by visual observations that were made using binoculars (i.e. Leica 10 × 42) and scopes (i.e. Swarowski 20-60 ×) by at least two experienced birders operating simultaneously. Visual observations were used only to detect if birds were moulting or not. Pictures and data were compared daily with those from the other watch points to delete possible double-counting of the same individuals according to the time and location of the birds passage as well as the individual moult features and sex. Only birds observed or photographed at closer range (< 100 m) were considered in the analysis, providing a random sample of the whole number of WMHs detected. We determined: (1) if harriers were moulting or not; (2) for a subsample of individuals, we were able to describe the moulting stage according to Ginn and Melville (1983). In this last subset of data, we recorded the number of primaries that the bird was moulting, if any. Therefore, we assigned a value from 1 to 10 according to the primary feathers moulting pattern of the species, being 1 = P1, 2 = P2 and so on (Baker 2016).
Data analysis
We used two different analyses. A Binary Logistic Regression Analysis (hereafter BLRA) (logit link function) (Boyce et al. 2002; Keating and Cherry 2004; Rushton et al. 2004) was used to test the factors influencing numbers of moulting individuals. For this we compared moulting and non-moulting birds (using both, data from visual observations and from pictures inspection), that in the BLRA was our dependent binary variable (1/0). The covariates were:
1. The Julian date calculated as the number of the day from the beginning of the year (January 1st being day number 1).
We tested the fitness of the model by means of the area under the curve of the Receiver Operating Characteristic (ROC) plot (Pearce and Ferrier 2000; Boyce et al. 2002; Fawcett 2006). This area provides a measure of discrimination ability, varying from 0.5 for a model with a discrimination ability no better than random, to 1.0 for a model with perfect discriminatory ability.
To verify the variation of moulting stage among the photographed individuals, we used a Linear Model (LM) using the number of moulting primaries as dependent variable (from 1 to 10). As covariates we used the same of the BLRA with the exclusion of the watchpoint. We tested the fitness of the model checking the R2 value.
For both, LM and BLRA, we made a stepwise model selection comparing the different models by the AICc value and choosing the ones with the lowest value. All statistical analyses were made with R open source software (R Development Core Team 2015).
Results
We observed a total of 3522 Western Marsh Harriers between the 13th August to 30th September. Among those, the individuals that were aged as adult birds were 1125 while juveniles were 413. From adult birds, 802 were positively identified as males and 323 as females, of which 19 were 2nd CY birds. Other 673 individuals were recorded under the category female/juvenile type. The remaining 1311 observed WMHs were undetermined. We took close pictures that allowed us to identify clear signs of moult from 107 individuals, while other 114 harriers did not show moult signs. From all these WMHs we obtained active moult schemes extracted from photos of 35 WMHs, 21 males and 14 females respectively. Another seven WMHs that recently suspended the moult of their primaries were confirmed but not included in the analyses because they weren't active moulting birds. We did not record any individuals with old feathers missing or with new pin feathers, neither individuals with new feathers at one-third of its whole length. The timing of the primary feathers moult varies slightly between males and females (Fig. 2). Females showed a more advanced moult stage on average than males, with 100% of the females with the inner 3rd primary already moulted unlike the 79% of the males. Similarly, different proportions between sexes have been recorded for the 4th, 5th, 6th and 7th primaries. However, the three outermost primaries, the 8th, the 9th and the 10th, remained unmoulted in both sexes (Fig. 3).
Figure
2.
Proportions of moulting WMH during autumn migration at the Strait of Messina. Moulting (Yes = light grey) and non-moulting (No = dark grey) birds across the season (11th August-30th September). (a) Males, (b) Females
The results of both, the BLRA and the LM (Table 1), indicate that the Julian date is the only relevant variable explaining the passage of moulting harriers in the study area, with higher numbers observed early in the season and with a more advanced moult later in the season (Table 1). Females were observed with more moulted primaries than males (Fig. 4). Moreover, 2nd CY harriers have a more advanced moult comparing to adults (Table 1).
Table
1.
Model selection for the BLRA and for the LM investigating factors influencing moult of migrating Western Marsh Harriers
Model
AICc
ΔAICc
AUC/R2
BLRA
Julian date (β = -0.04 ± 0.01, p < 0.01)
288.6
0
0.61
Julian date, Julian date squared, Sex, Age, Watchpoint
293.6
5
0.64
LM
Julian date (β = 0.1 ± 0.03, p < 0.05), Sex (Males, β = -1 ± 0.4, p < 0.05), Age (Adults, β = -1.7 ± 0.8, p < 0.05)
22.39
0
0.48
Julian date, Julian date squared, Sex, Age
24.12
1.73
0.49
Parameters estimates ± standard errors are shown together with p values for the best models. The best models are shown in italics, below the full models are reported
Our results suggest that adult WMHs start their autumn migration once the last primary to be moulted (maximum until the P7) was already expelled, and the previous primaries are already growing. Growing stages of those feathers could be visible during the lapse of migration that it takes for feathers to grow until their complete development. Therefore, during post-breeding migration only growing new flight feathers and moulted new ones were recorded, but no lacking feathers. We detected the presence of primary feathers unmoulted, already moulted, and growing at medium/late moulting stages. Not a single individual showed the lack of a primary or an immediate growing stage such as 1 or 2 (Ginn and Melville 1983). This indicates that replacing the next old feathers in the sequence is unlikely to occur during migration but in the winter quarters.
The lack of feathers might imply a less efficient flight with a disproportional increase of the energy required for flapping (Pennycuick 2008). Therefore, it is possible that adult WMHs which belong to the same sex group with a higher number of primary feathers involved in their moult should come from breeding grounds farther away than individuals showing a less extended ongoing moult. Moreover, our data suggests that there is a relationship between the extension and timing of the suspended moult and the timing of the migration itself with a difference between males and females. We found adult females showed an averaged more extended moult than the adult males and a later timing of migration as well (Agostini and Panuccio 2010; Agostini et al. 2017). This confirms that adult females start moulting in the breeding season before the males. However, they suspend that moult later on average as well (Cramp and Simmons 1980; Kjellén 1992).
Conclusion
If we considered "active moult" as the lapse of time during which a bird replaces feathers rather than the time spent by a bird on feathers growth, we might consider that active moult of WMHs ends at the breeding ground immediately after shedding the last primary to be replaced and just before starting the post-breeding migration. So, during migration, only growing feathers take place, being the extension of the suspended moult already defined at the breeding ground just before leaving. This strategy might be evolved to minimize the effect of moulting during migration. Such movements are energetically demanding and harriers are known to be raptors that largely use powered-flapping flight over long distances (Spaar and Bruderer 1997; Panuccio et al. 2013, 2016; Agostini et al. 2015, 2017).
Authors' contributions
JR made the study design, collected data on moulting, prepared the dataset and wrote the first draft of the manuscript. MP coordinated the fieldwork, made the analysis and wrote the paper. Both authors read and approved the final manuscript.
Acknowledgements
We thank Giacomo Dell'Omo and Ornis italica for his support during the fieldwork. We are grateful to all the people who collected data at the Strait of Messina in autumn 2016 and in particular: Alberto Pastorino, Michele Cento, Gianpasquale Chiatante, Nicolantonio Agostini, Martina Zambon, Stiven Kocijančič, Giuseppe Cicero. We would also like to acknowledge Kylynn Clare for her contributions to the editing of this paper. We wish to thank MEDRAPTORS (www.raptormigration.org), a network of ornithologists involved in the research and the protection of migrating birds of prey through specific projects and observation camps.
Competing interests
The authors declare that they have no competing interests.
Availability of data and materials
The datasets used in the present study are available from the corresponding author on request.
Baumgardt JA, Goldberg CS, Reese KP, Connelly JW, Musil DD, Garton EO, Waits LP. A method for estimating population sex ratio for sage-grouse using noninvasive genetic samples. Mol Ecol Resour. 2013;13:393–402.
BirdLife International. Species factsheet: Egretta eulophotes. 2014. [].
Bonin A, Bellemain E, Bronken Eidesen P, Pompanon F, Brochmann C, Taberlet P. How to track and assess genotyping errors in population genetics studies. Mol Ecol. 2004;13:3261–73.
Brooks R, Williamson J, Hensley A, Butler E, Touchton G, Smith E. Buccal cells as a source of DNA for comparative animal genomic analysis. Biotech Lett. 2003;25:451–4.
Broquet T, Petit E. Quantifying genotyping errors in noninvasive population genetics. Mol Ecol. 2004;13:3601–8.
Brown MB, Brown CR. Blood sampling reduces annual survival in cliff swallows (Petrochelidon pyrrhonota). Auk. 2009;126:853–61.
Bush KL, Vinsky MD, Aldridge CL, Paszkowski CA. A comparison of sample types varying in invasiveness for use in DNA sex determination in an endangered population of greater Sage-Grouse (Centrocercus uropihasianus). Conserv Genet. 2005;6:867–70.
Costantini V, Guaricci AC, Laricchiuta P, Rausa F, Lacalandra GM. DNA sexing in Humboldt Penguins (Spheniscus humboldti) from feather samples. Anim Reprod Sci. 2008;106:162–7.
Criscuolo F. Does blood sampling during incubation induce nest desertion in the female common eider Somateria mollissima. Mar Ornithol. 2001;29:47–50.
Dai Y, Zhou X, Fang W, Lin Q, Chen X. Development and cross-species transferability of 23 microsatellite markers from the vulnerable Chinese Egret (Egretta eulophotes) using 454 sequencing. Conserv Genet Resour. 2013;5:1035–8.
Deagle BE, Eveson JP, Jarman SN. Quantification of damage in DNA recovered from highly degraded samples—a case study on DNA in faeces. Front Zool. 2006;3:11.
Egloff C, Labrosse A, Hebert C, Crump D. A nondestructive method for obtaining maternal DNA from avian eggshells and its application to embryonic viability determination in herring gulls (Larus argentatus). Mol Ecol Resour. 2009;9:19–27.
Gagneux P, Boesch C, Woodruff DS. Microsatellite scoring errors associated with noninvasive genotyping based on nuclear DNA amplified from shed hair. Mol Ecol. 1997;6:861–8.
Gebhardt KJ, Waits LP. High error rates for avian molecular sex identification primer sets applied to molted feathers. J Field Ornithol. 2008;79:286–92.
Handel CM, Pajot LM, Talbot SL, Sage GK. Use of buccal swabs for sampling DNA from nestling and adult birds. Wildl Soc Bull. 2006;34:1094–100.
Horvâth MB, Martìnez CB, Negro JJ, Kalmâr L, Godoy JA. An overlooked DNA source for non-invasive genetic analysis in birds. J Avian Biol. 2005;36:84–8.
Huang X, Zhou X, Chen M, Fang W, Chen X. Isolation and characterization of microsatellite loci in vulnerable Chinese egret (Egretta eulophotes: Aves). Conserv Genet. 2010;11:1211–4.
Huang X, Zhou X, Lin Q, Peng Z, Fang W, Chen X. A novel multiplex PCR assay for species identification in the Chinese Egret (Egretta eulophotes) and Little Egret (E. garzetta). Conserv Genet Resour. 2012a;4:31–3.
Huang X, Zhou X, Lin Q, Fang W, Chen X. An efficient molecular sexing of the vulnerable Chinese egret (Egretta eulophotes) from faeces samples. Conserv Genet Resour. 2012b;4:391–3.
Huang X, Zhou X, Lin Q, Fang W, Chen X. PCR-RFLP technique for species identification of molted feathers in six species of co-occurring Ardeids. Conserv Genet Resour. 2013;5:817–9.
Idaghdour Y, Broderick D, Korrida A. Faeces as a source of DNA for molecular studies in a threatened population of great bustards. Conserv Genet. 2003;4:789–92.
Isabel MC, Del Lama Sn. Molted feathers as a source of DNA for genetic studies in waterbird populations. Waterbirds. 2009;32:322–9.
Johansson MP, Mcmahon BJ, Höglund J, Segelbacher G. Amplification success of multilocus genotypes from feathers found in the field compared with feathers obtained from shot birds. Ibis. 2012;154:15–20.
Kalinowski ST, Taper ML, Marshall TC. Revising how the computer program CERVUS accommodates genotyping error increases success in paternity assignment. Mol Ecol. 2007;16:1099–106.
Kang JH, Kim IK, Lee H, Rhim SJ. Distribution and Breeding Status of Chinese Egret Egretta eulophotes in South Korea. J Anim Vet Adv. 2013;12:618–20.
Litvinenko NM, Shibaev YV. Importance of Furugelm Island in the Sea of Japan for wetland birds: the first record of a breeding colony of the Chinese egret Egretta eulophotes. Oryx. 2000;34:335–7.
Martín-Gálvez D, Peralta SJ, Dawson D, Martín PA, Martínez BM, Burke T, Soler JJ. DNA sampling from eggshell swabbing is widely applicable in wild bird populations as demonstrated in 23 species. Mol Ecol Resour. 2011;11:481–93.
Matysioková B, Remeš V. The importance of having a partner: male help releases females from time limitation during incubation in birds. Front Zool. 2014;11:24.
Miquel C, Bellemain E, Poillot C, Bessiere J, Durand A, Taberlet P. Quality indexes to assess the reliability of genotypes in studies using noninvasive sampling and multiple-tube approach. Mol Ecol Notes. 2006;6:985–8.
Pompanon F, Bonin A, Bellemain E, Taberlet P. Genotyping errors: causes, consequences and solutions. Nat Rev Genet. 2005;6:846–7.
Regnaut S, Lucas FS, Fumagalli L. DNA degradation in avian faecal samples and feasibility of non-invasive genetic studies of threatened capercaillie populations. Conserv Genet. 2006;7:449–53.
Renan S, Speyer E, Shahar N, Gueta T, Templeton AR, Bar D. A factorial design experiment as a pilot study for noninvasive genetic sampling. Mol Ecol Resour. 2012;12:1040–7.
Rousset F. GENEPOP'007: a complete re-implementation of the genepop software for Windows and Linux. Mol Ecol Resour. 2008;8:103–6.
Sacchi P, Soglia D, Maione S, Meneguz G, Campora M, Rasero R. A non-invasive test for sex identification in Short-toed Eagle (Circaetus gallicus). Mol Cell Probe. 2004;18:193–6.
Schmaltz G, Somers CM, Sharma P, Quinn JS. Non-destructive sampling of maternal DNA from the external shell of bird eggs. Conserv Genet. 2006;7:543–9.
Shen Z, Qu W, Wang W, Lu Y, Wu Y, Li Z, Zhang C. MPprimer: a program for reliable multiplex PCR primer design. BMC Bioinformatics. 2010;11:143.
Skutch AF. The incubation patterns of birds. Ibis. 1957;99:69–93.
Taberlet P, Bouvet J. A single plucked feather as a source of DNA for bird genetic studies. Auk. 1991;108:959–60.
Taberlet P, Griffin S, Goossens B, Questiau S, Manceau V, Escaravage N, Bouvet J. Reliable genotyping of samples with very low DNA quantities using PCR. Nucleic Acids Res. 1996;24:3189–94.
Taberlet P, Waits LP, Luikart G. Noninvasive genetic sampling: look before you leap. Trends Ecol Evol. 1999;14:323–7.
Van Oosterhout C, Hutchinson WF, Wills DP, Shipley P. MICRO-CHECKER: software for identifying and correcting genotyping errors in microsatellite data. Mol Ecol Notes. 2004;4:535–8.
Wang Z, Zhou X, Lin Q, Fang W, Chen X. New primers for sex identification in the Chinese Egret and other ardeid species. Mol Ecol Resour. 2011;11:176–9.
Wang Z, Zhou X, Lin Q, Fang W, Chen X. Characterization, polymorphism and selection of major histocompatibility complex (MHC) DAB genes in vulnerable Chinese egret (Egretta eulophotes). PLoS One. 2013;8:e74185.
Wellbrock AH, Bauch C, Rozman J, Witte K. Buccal swabs as a reliable source of DNA for sexing young and adult Common Swifts (Apus apus). J Ornithol. 2012;153:991–4.
Yannic G, Sermier R, Aebischer A, Gavrilo MV, Gilg O, Miljeteig C, Broquet T. Description of microsatellite markers and genotyping performances using feathers and buccal swabs for the ivory gull (Pagophila eburnea). Mol Ecol Resour. 2011;11:877–89.
Zhou X, Wang Y, Chen X, Lin Q, Fang W, Wei D. A set of primer pairs for amplifying the complete mitochondrial DNA of endangered Chinese egret (Aves, Ardeidae, Egretta eulophotes). Mol Ecol Resour. 2008;8:412–4.
Zhou X, Fang W, Chen X. Mitochondrial DNA diversity of the vulnerable Chinese Egret (Egretta eulophotes) from China. J Ornithol. 2010;151:409–14.
Zhou X, Lin Q, Fang W, Chen X. The complete mitochondrial genomes of sixteen ardeid birds revealing the evolutionary process of the gene rearrangements. BMC Genom. 2014;15:573.
Li, J., Liu, D., Li, W. et al. Moult-breeding trade-offs in the Crested Ibis (Nipponia nippon) could have management applications. Avian Research, 2025, 16(2): 100250.
DOI:10.1016/j.avrs.2025.100250
2.
Adekola, O.E., Allan, D.G., Bernitz, Z. et al. Extent and symmetry of tail moult in Amur Falcons. Journal of Ornithology, 2021, 162(3): 655-667.
DOI:10.1007/s10336-021-01874-0
3.
Nadal, J., Ponz, C., Margalida, A. The end of primary moult as an indicator of global warming effects in the Red-legged Partridge Alectoris rufa, a medium sized, sedentary species. Ecological Indicators, 2021.
DOI:10.1016/j.ecolind.2020.107287
4.
Agostini, N., Chiatante, G., Dell’Omo, G. et al. Differential autumn migration between sex and age groups in the Western marsh harrier: a longitudinal pattern analysis. Ethology Ecology and Evolution, 2021, 33(1): 73-82.
DOI:10.1080/03949370.2020.1820581
5.
Bounas, A.. Premigratory moult in the Lesser Kestrel Falco naumanni. Avocetta, 2019, 43(1): 49-54.
DOI:10.30456/AVO.2019106
Key Laboratory of the Ministry of Education for Coast and Wetland Ecosystems, School of Life Sciences, College of the Environment and Ecology, Xiamen University, Xiamen 361102, China
Table
1.
Model selection for the BLRA and for the LM investigating factors influencing moult of migrating Western Marsh Harriers
Model
AICc
ΔAICc
AUC/R2
BLRA
Julian date (β = -0.04 ± 0.01, p < 0.01)
288.6
0
0.61
Julian date, Julian date squared, Sex, Age, Watchpoint
293.6
5
0.64
LM
Julian date (β = 0.1 ± 0.03, p < 0.05), Sex (Males, β = -1 ± 0.4, p < 0.05), Age (Adults, β = -1.7 ± 0.8, p < 0.05)
22.39
0
0.48
Julian date, Julian date squared, Sex, Age
24.12
1.73
0.49
Parameters estimates ± standard errors are shown together with p values for the best models. The best models are shown in italics, below the full models are reported


 DownLoad:
DownLoad:








 Email Alerts
Email Alerts RSS Feeds
RSS Feeds