Bei An, Lixun Zhang, Yutao Wang, Sen Song. 2020: Comparative phylogeography of two sister species of snowcock: impacts of species-specific altitude preference and life history. Avian Research, 11(1): 1. DOI: 10.1186/s40657-019-0187-0
Citation:
Bei An, Lixun Zhang, Yutao Wang, Sen Song. 2020: Comparative phylogeography of two sister species of snowcock: impacts of species-specific altitude preference and life history. Avian Research, 11(1): 1. DOI: 10.1186/s40657-019-0187-0
Bei An, Lixun Zhang, Yutao Wang, Sen Song. 2020: Comparative phylogeography of two sister species of snowcock: impacts of species-specific altitude preference and life history. Avian Research, 11(1): 1. DOI: 10.1186/s40657-019-0187-0
Citation:
Bei An, Lixun Zhang, Yutao Wang, Sen Song. 2020: Comparative phylogeography of two sister species of snowcock: impacts of species-specific altitude preference and life history. Avian Research, 11(1): 1. DOI: 10.1186/s40657-019-0187-0
School of Basic Medicine Sciences, Lanzhou University, Lanzhou 730000, China
2.
School of Life Sciences, Lanzhou University, Lanzhou 730000, China
3.
Gansu Key Laboratory of Biomonitoring and Bioremediation for Environmental Pollution, Lanzhou University, Lanzhou 730000, China
4.
College of Life and Geographic Sciences, Kashi University, Kashi 844000, China
5.
The Key Laboratory of Ecology and Biological Resources in Yarkand Oasis at Colleges & Universities under the Department of Education of Xinjiang Uygur Autonomous Region, Kashi University, Kashi 844000, China
Funds:
the Strategic Priority Research Program of Chinese Academy of SciencesXDA2010010103
National Natural Science Foundation of China31372195
National Natural Science Foundation of China31772436
the Open Foundation of Research Institute of Qilian Mountains, Lanzhou University
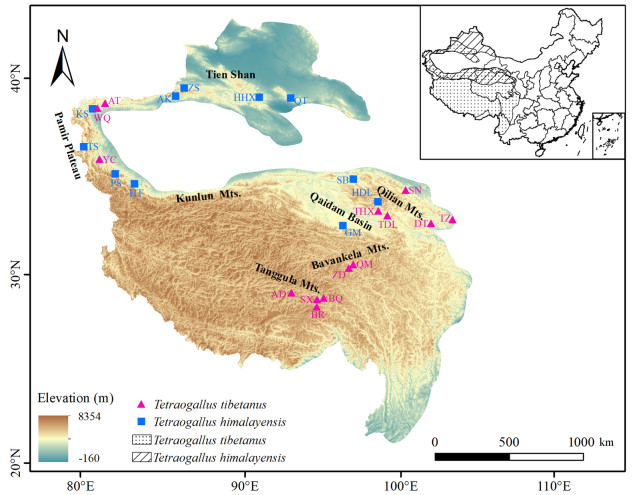
Phylogeographical patterns and population dynamics are usually interpreted by environmental disturbances and geographic barriers of the past. However, sister species may exhibit disparate patterns of genetic structures and population dynamics due to their habitat preference and altitude segregation. In this study, we tested how species-specific altitude habitat affected phylogeographical patterns in two sister snowcock species, Tibetan (Tetraogallus tibetanus) and Himalayan Snowcocks (T. himalayensis).
Methods
A panel of seven microsatellite loci and a fragment of Mitochondrial DNA Control Region were used to investigate genetic structures and population dynamics in hope of revealing the underlying evolutionary processes through the identification of possible past demographic events.
Results
Our results suggest that T. himalayensis showed a significant phylogeographical signal in mtDNA (FST = 0.66, p < 0.001) and microsatellite (FST = 0.11, p < 0.001) data and is stable during the glacial-interglacial cycles in the Pleistocene and followed demographic contraction until 0.003 million years (Mys) ago. The phylogeographical signal of T. tibetanus is lower than the level of genetic difference among populations in mtDNA (FST = 0.41, p < 0.001) and microsatellite (FST = 0.09, p < 0.001) data, likely benefiting from stable habitats over a long period of time. T. tibetanus has been experiencing expansion since 0.09 Mys ago. However, an abnormally haplotype H9 from T. himalayensis clustering with T. tibetanus was spotted.
Conclusion
Our results indicate that differences in habitat preference and altitude specialities were reflected in the genetic structure patterns and population dynamics of these two species. These dissimilarities in life history traits might have affected the dispersal and survival abilities of these two species differently during environmental fluctuations. The results of this study also enriched our knowledge on population differentiation and connectivity in high altitude mountain ecosystems.
Mothers have a variety of ways to manipulate the composition and/or environment of their eggs, including, but not limited to: hatching asynchrony (Clark and Wilson 1981; Magrath 1990), yolk hormones (Groothuis et al. 2005), sex allocation (Pike and Petrie 2003), egg size (Krist 2011), incubation temperature (DuRant et al. 2009, 2011), and carotenoids (Royle et al. 2001; Blount et al. 2002). Assessing how these factors influence the growth and survival of their offspring, however, first requires being able to accurately match hatchlings to their respective eggs. This can be a challenging endeavor for many bird species.
Frequent nest checks can be employed to directly observe the individual hatching of eggs, although increased disturbance at the nest may lead to higher rates of abandonment or predation. Even the most frequent nest checks, however, can result in finding two hatchlings within the same nest. Because egg and hatchling mass are significantly correlated in most bird species (Williams 1994; Deeming and Birchard 2007), simply assigning the heaviest hatchling to the heaviest egg (i.e., relative mass assignment; RMA) in these situations could be tempting, as RMA would increase sample sizes and reduce the number of nest checks required.
Here I present RMA as a new technique, at least to my knowledge, that may be useful in studies of maternal effects, and determine the accuracy of RMA by applying the method retrospectively to a dataset where Common Grackle (Quiscalus quiscula) hatchlings were assigned to eggs unambiguously. Common Grackles are an appropriate species in which to test the utility of RMA because hatchling mass is correlated with egg mass (Howe 1976), egg size is variable both within and among clutches (Howe 1976; Maddox and Weatherhead 2008), and 2‒4 eggs within a given clutch can hatch at approximately the same time (Howe 1976; Maddox and Weatherhead 2008).
Methods
I used an incubator with individual compartments to unambiguously assign hatchlings to their respective eggs as part of a larger study investigating maternal effects in Common Grackles. Detailed descriptions of the study site and field methods can be found in Maddox and Weatherhead(2008, 2009, 2012). Briefly, I found nests under construction and monitored them daily until fledging. On the day each egg was laid, I measured its mass (±0.1 g) and numbered it sequentially with a nontoxic felt-tip marker. When I observed at least one egg pipping from an individual nest during my daily nest checks, I collected all the eggs from the clutch and placed each egg in an individual compartment within the incubator. In total, eggs from 133 nests were treated as such. Artificial eggs of similar size and appearance were placed in nests to limit female abandonment. Upon hatching I measured the mass (±0.1 g), wing chord (±0.1 mm) and tarsus (±0.1 mm) of each hatchling before returning it to its respective nest.
When more than one egg hatched between checks of the incubator or approximately every 1‒2 h, I used RMA to assign hatchlings to their respective eggs by rank-ordering the mass of hatchlings and their candidate eggs separately and assigning hatchlings to eggs of the same rank (i.e., 1-1, 2-2, etc.). Because I knew the source egg in each case, I could evaluate the error rate of RMA. Typically two eggs from a given clutch hatched between checks but finding 3‒4 hatchlings was not uncommon. This is essentially identical to what occurs in the field with frequent nest checks.
Results
In 78 of the 133 clutches examined, two eggs hatched between consecutive checks of the incubator. RMA correctly matched 122 (78.2%) hatchlings to their respective eggs, whereas 14 (9.0%) hatchlings were incorrectly assigned. RMA could not be applied to 20 (12.8%) hatchlings because the mass of the two hatchlings or the two candidate eggs was identical. For those egg-hatchling dyads that were assigned correctly, paired differences of egg and hatchling masses averaged 0.40 g ± 0.03 SE (range 0.1-0.9 g, n = 61) and 0.38 g ± 0.04 SE (range 0.1-1.4 g, n = 61), respectively. For the 14 egg-hatchling dyads that were assigned incorrectly, paired average differences of egg (0.23 g ± 0.07 SE; range 0.1-0.6 g) and hatchling (0.21 g ± 0.04 SE; range 0.1-0.4 g) masses were almost half that of correctly assigned dyads. Excluding those hatchlings that could not be assigned, the error rate of RMA when applied to two hatchlings was 10.3% (14/136).
In 50 of the 133 clutches, three eggs hatched between consecutive incubator checks. RMA correctly matched 66 (44%) hatchlings to their respective eggs, whereas 5 clutches (10%) had at least one incorrectly assigned hatchling. Twenty-three (46%) nests contained at least one hatchling that could not be assigned to an egg for the reasons indicated above. Differences in egg mass of correct assignments averaged 0.25 ± 0.03 SE, whereas wrong assignments averaged 0.26 ± 0.02 SE. Excluding unassigned hatchlings, the error rate of RMA was 7.0% (5/71).
In the remaining nine clutches, 4 eggs within the same nest hatched between incubator checks. In one (14%) clutch, all 4 hatchlings were assigned correctly to their eggs. In another (14%) clutch, at least one hatchling was incorrectly assigned. In 7 (78%) clutches, at least one hatchling could not be assigned to an egg for the reasons indicated above. Excluding unassigned hatchlings, the error rate of RMA was 50% (1/2).
Discussion
RMA was largely successful at correctly assigning hatchlings to their respective eggs. The ability of RMA to assign eggs, however, was highly dependent on the number of hatchlings present between checks. When applied to ≥3 unknown egg-hatchling dyads the percentage of hatchlings that RMA was unable to assign was substantial, essentially eliminating its use in those situations. Consequently, I focus the remaining discussion to those cases where only two hatchlings were found between checks. With only two hatchlings present, RMA may prove to be a useful technique in some species, but its widespread adoption will likely be restricted by several shortcomings.
The accuracy of RMA is likely to vary directly with the extent that egg and hatchling mass are correlated. RMA will almost certainly perform below acceptable levels when egg mass explains little of the variation in hatchling mass. Indeed, those dyads that were incorrectly assigned generally had greater residuals than correctly assigned dyads (Fig. 1). Given that egg mass explained 82% of the variation in hatchling mass in Common Grackles—a high value for most bird species (Williams 1994)—and yet still incurred a 10% error rate, RMA should be limited to species in which egg and hatchling mass are highly correlated.
Figure
1.
Relative mass assignment performed better when the difference in egg mass was greater with incorrectly assigned eggs generally having greater residuals than correctly assigned dyads. Colors indicate correct (green), incorrect (red), or unknown (blue) assignments of egg-hatchling dyads
Restricting the application of RMA by a pre-established minimum difference in egg or hatchling mass may improve the error rate, because the pair-wise difference in mass was smaller for incorrectly assigned egg-hatchling dyads than those that were correctly assigned (Fig. 1). Limiting RMA in the current dataset to only eggs that differed in mass by > 0.1 g, reduced the error rate in half to 5%, but doubled the percentage of hatchlings that were unable to be assigned from 13 to 27%. Limiting the assignment of hatchlings had no discernible effect.
A potential pitfall of RMA not accounted for in the present study is the ability of parents to feed hatchlings between nest checks. Given the small difference in mass between nest mates, any amount of food given differentially to hatchlings could alter the mass difference relative to their nest mate and thus potentially result in a wrong assignment. This may prove negligible in many species, especially in species where males provide little parental care, as it is likely that females would still be brooding hatchlings and/or incubating eggs. Common Grackle hatchlings, however, started begging for food immediately after hatching, and I did occasionally observe males feeding hatchlings. Consequently, the error rate I report here is likely a best-case scenario.
One modification to RMA that may improve its accuracy that the current study was unable to assess is to measure egg mass directly before hatching. Given that egg mass decreases during incubation due to water loss as a result of embryonic development (Ar and Rahn 1980), egg mass obtained after this mass loss in addition to or instead of fresh egg mass may decrease the number of incorrect assignments. Future studies will need to verify if this modification would markedly increase the accuracy of RMA.
Conclusions
RMA may prove to be a valuable tool for studies that require known egg-hatchling dyads, although its accuracy should first be tested on the species in which it will ultimately be applied. Its successful application will likely be limited to those species where egg and hatchling mass are highly correlated or within-clutch egg-size variation is large. Researchers should be cognizant of the fact that RMA is a one-tailed error—egg-size effects will always be inflated—and thus could potentially result in erroneously reporting significant effects when in fact they do not exist.
Acknowledgements
I am indebted to the Shoemaker family for allowing me to work on their land and to the many people who contributed to this project over the years, especially K. Kopatz, C. Johnson, D. Kikuchi, and P. Wesley. Funding was provided by the University of Illinois, Program in Ecology, Evolution and Conservation, and the American Ornithologists' Union Research Award. L.S. Johnson provided useful comments on an earlier version of the manuscript. I was supported by an NSF International Fellowship OISE-1159178 while preparing an earlier version of this manuscript.
Competing interests
The author declares that he has no competing interests.
Ethical standards
All work described here was approved by the Institutional Animal Care and Use Committee (IACUC) at the University of Illinois (protocol #05006).
Adeli, Ma M, Hairoula, Wang Z. The ecological habits and characteristics of snowcock in the southern part of Bogeda, Tianshan Mountains. Arid Zone Res. 1997;14:84-7 (in Chinese).
An B, Zhang LX, Browne S, Liu NF, Ruan LZ, Song S. Phylogeography of tibetan snowcock (Tetraogallus tibetanus) in Qinghai-Tibetan plateau. Mol Phylogenet Evol. 2009;50:526-33.
An B, Zhang L, Liu N, Wang Y. Refugia persistence of Qinghai-Tibetan Plateau by the cold-tolerant bird Tetraogallus tibetanus (Galliformes: phasianidae). PLoS ONE. 2015;10:e0121118.
Ashcroft MB, Gollan JR, Warton DI, Ramp D. A novel approach to quantify and locate potential microrefugia using topoclimate, climate stability, and isolation from the matrix. Glob Change Biol. 2012;18:1866-79.
Bandelt HJ, Forster P, Rohl A. Median-joining networks for inferring intraspecific phylogenies. Mol Biol Evol. 1999;16:37-48.
Benzécri JP. L'analyse Des Données: Tome 2, L'Analyse Des Correspondances. Paris, France: Dunod; 1973.
Bermingham E, Moritz C. Comparative phylogeography: concepts and applications. Mol Ecol. 1998;7:367-9.
Bianki VL. Review of the species of genus Tetraogallus Gray (1898). In: Proceedings of Museum Emperor's Academy of Sciences, St. Petersburg. 2001;3:113-23. (in Russian).
Correa Ribeiro P, Lemos-Filho JP, Oliveira BRS, Lovato MB, Heuertz M. Species-specific phylogeographical patterns and pleistocene east-west divergence in annona (Annonaceae) in the Brazilian cerrado. Bot J Linn Soc. 2016;181:21-36.
Darriba D, Taboada GL, Doallo R, Posada D. Jmodeltest 2: more models, new heuristics and parallel computing. Nat Methods. 2012;9:772.
Drummond AJ, Rambaut A. BEAST: bayesian evolutionary analysis by sampling trees. BMC Evol Biol. 2007;7:214.
Earl DA, vonHoldt BM. STRUCTURE HARVESTER: a website and program for visualizing STRUCTURE output and implementing the Evanno method. Conserv Genet Res. 2012;4:359-61.
Excoffier L, Lischer HEL. ARLEQUIN suite ver 3.5: a new series of programs to perform population genetics analyses under Linux and Windows. Mol Ecol Resour. 2010;10:564-7.
Fang F, Chen J, Jiang LY, Chen R, Qiao GX. Biological traits yield divergent phylogeographical patterns between two aphids living on the same host plants. J Biogeogr. 2017;44:1-13.
Fu YX. Statistical tests of neutrality of mutations against population growth, hitchhiking and background selection. Genetics. 1997;147:915.
Goudet J. FSTAT, version 2.9.3. A program to estimate and test gene diversities and fixation indices. Lausanne: Lausanne University; 2001.
Hubisz MJ, Falush D, Stephens M, Pritchard JK. Inferring weak population structure with the assistance of sample group information. Mol Ecol Resour. 2009;9:1322-32.
Janecka JE, Zhang Y, Li D, Munkhtsog B, Bayaraa M, Galsandorj N, et al. Range-wide Snow Leopard phylogeography supports three subspecies. J Hered. 2017;108:597-607.
Kennedy JD, Price TD. Ecological limits on diversification of the Himalayan core Corvoidea. Evolution. 2012;66:2599-613.
Lei F, Qu Y, Song G. Species diversification and phylogeographical patterns of birds in response to the uplift of the Qinghai-Tibet Plateau and Quaternary glaciations. Curr Zool. 2014;60:149-61.
Li J, Xiao L, Lu Z. Challenges of snow leopard conservation in China. Sci China Life Sci. 2016;59:637-9.
Librado PRJ. DnaSP v5: a software for comprehensive analysis of DNA polymorphism data. Bioinformatics. 2009;25:1451-2.
Liu N. Isolating mechanism between Tibetan snowcock (Tetraogallus tibetanus) and Himalayan snowcock (Tetraogallu himalayensis). The 65th anniversary meeting of the China Zoological Society, Beijing. 1999. p. 235-46.
Ma M, Xu F, Zhang T, Tuergan M, Ding P, Chen Y. The Tetraogallu himalayensis and Tetraogallus tibetanus mixed in the same area of the Altun-Kunlun Montains. Thesis volume in the channel-intercoastal science session. 2013.
McDonald JH, Kreitman M. Adaptive evolution at the Adh locus in Drosophila. Nature. 1991;351:652-4.
Namgail T. Winter birds of the Gya-Miru Wildlife Sanctuary, Ladakh, Jammu and Kashmir, India. Indian Birds. 2005;1:26-8.
Price TD, Hooper DM, Buchanan CD, Johansson US, Tietze DT, Alström P, et al. Niche filling slows the diversification of Himalayan songbirds. Nature. 2014;509:222-5.
Pritchard JK, Stephens M, Donnelly P. Inference of population structure using multilocus genotype data. Genetics. 2000;155:945-59.
Prum RO, Berv JS, Dornburg A, Field DJ, Townsend JP, Lemmon EM, et al. A comprehensive phylogeny of birds (Aves) using targeted next-generation DNA sequencing. Nature. 2015;526:569-73.
Qu Y, Lei F, Zhang R, Lu X. Comparative phylogeography of five avian species: implications for pleistocene evolutionary history in the Qinghai-Tibetan Plateau. Mol Ecol. 2009;19:338-51.
Qu Y, Zhang R, Quan Q, Song G, Li S, Lei F. Incomplete lineage sorting or secondary admixture: disentangling historical divergence from recent gene flow in the Vinous-throated Parrotbill (Paradoxornis webbianus). Mol Ecol. 2012;21:6117-33.
Rogers AR, Harpending H. Population growth makes waves in the distribution of pairwise genetic differences. Mol Biol Evol. 1992;9:552-69.
Ruan L, Zhang L, Wen L, Sun Q, Liu N. Phylogeny and molecular evolution of Tetraogallus in China. Biochem Genet. 2005;43:507-18.
Shen X, Wang J. Systematics, geographical distribution, and ecology of Tetraogallus in China. Chin J Zool. 1963;5:186-92.
Tajima F. Statistical-method for testing the neutral mutation hypothesis by DNA polymorphism. Genetics. 1989;123:585-95.
Wang X, Qu J, Liu N, Bao X, Song S. Limited gene flow and partial isolation phylogeography of Himalayan snowcock, Tetraogallus himalayensis, based on part mitochondrial D-loop sequences. Curr Zool. 2011;57:758-67.
Weir JT, Schluter D. Calibrating the avian molecular clock. Mol Ecol. 2008;17:2321-8.
Zhang R, Song G, Qu Y, Alström P, Ramos R, Xing X, et al. Comparative phylogeography of two widespread magpies: importance of habitat preference and breeding behavior on genetic structure in China. Mol Phylogenet Evol. 2012;65:562-72.
Zhang H, Zhang ML, Sanderson SC. Retreating or standing: responses of forest species and steppe species to climate change in arid Eastern Central Asia. PLoS ONE. 2013;8:e61954.
Zhang L, Shu M, Zhao C. The Tetraogallus tibetanus and T. himalayensis coexisted. China Nat. 2015;2:50-3.
Zhang M, Mei J, Zhang Z, Wang J, Xu X. Be exposure ages obtained from quaternary glacial landforms on the Tibetan Plateau and in the surrounding area. Acta Geol Sin-Engl. 2018;92:366-80.
Zheng Z. Fauna Sinica: Aves, vol. 4. Beijing: Science Press; 1978. p. 51-8.


 DownLoad:
DownLoad:





 Email Alerts
Email Alerts RSS Feeds
RSS Feeds