
Figure
1.
Map of the study area
| Citation: |
Johannes Fritz, Barbara Eberhard, Corinna Esterer, Bernhard Goenner, Daniela Trobe, Markus Unsoeld, Bernhard Voelkl, Helena Wehner, Alexandra Scope. 2020: Biologging is suspect to cause corneal opacity in two populations of wild living Northern Bald Ibises (Geronticus eremita). Avian Research, 11(1): 38. DOI: 10.1186/s40657-020-00223-8

|
In this paper, we present evidence that biologging is strongly correlated with eye irritation, with sometimes severely impairing effects. A migratory population of the Northern Bald Ibis (Geronticus eremita, NBI) is reintroduced in Europe, in course of a LIFE + project. Since 2014, all individuals have been equipped with GPS-devices. Remote monitoring allows the implementation of focussed measures against major mortality causes.
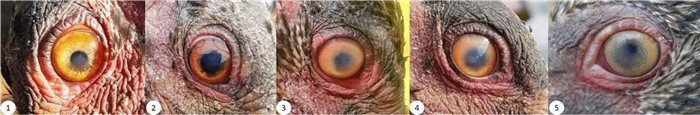
Initially all birds carried battery-powered devices, fixed on the lower back of the birds. Since 2016 an increasing amount of birds has been equipped with solar-powered devices, fixed on the upper back, the more sun-exposed position. In 2016, we observed opacity in the cornea of one eye (unilateral corneal opacity; UCO) during a regular health monitoring for the first time.
By 2018, a total of 25 birds were affected by UCO, with varying intensity up to blindness. Clinical examination of the birds revealed no clear cause for the symptoms. However, only birds carrying a device on the upper back were affected (2017 up to 70% of this group). In contrast, none of the birds carrying devices on the lower back ever showed UCO symptoms. This unexpected relationship between tagging and UCO was discovered in 2017. After we took countermeasures by removing the device or repositioning it on the lower back, we observed an immediate reduction of the incidence rate without any new cases reported since January 2019. NBI roost with their head on the back, one eye closely placed to the device if it was positioned on the upper back. Thus, we conclude that the most parsimonious explanation for the symptomatology is either a repetitive slight temperature rise in the corneal tissue due to electromagnetic radiation by the GSM module of the device or a repetitive slight mechanical irritation of the corneal surface. Concrete evidence is missing so far. Meanwhile, cases of UCO were found in another NBI population.
Our observations indicate that further research in the fast-growing field of biologging is urgently needed. The findings question the positioning of devices on the upper back in birds roosting with the head on the back.
Inland and coastal fisheries and waterbirds are inextricably linked (Green and Elmberg 2014). Waterbirds around the world are declining as a result of habitat loss, disturbance or alterations in habitat quality sometimes caused by fishing activities (Butchart et al. 2010; Aarif et al. 2014; Green and Elmberg 2014; Bundy et al. 2017). Shrinking wetlands have reduced waterbird wintering areas and have caused changes in diet and habitat use patterns in many waterbird assemblages.
Fish species composition, for example, often significantly affects habitat use by waterbirds due to their abundance in selected microhabitats (De Nie 1995; Warke and Day 1995; Elmberg et al. 2010; Kloskowski et al. 2010; Russell et al. 2014). Lakes supporting large fish may be used by an assemblage of diving, piscivorous waterbirds (Paszkowski and Tonn 2000). Fish-eating waterbirds often profit from local increases in fish populations (Lammens 1999), whereas negative effects of fish on waterbird distribution have been documented and attributed to exploitative competition (Hurlbert et al. 1986; Van Eerden et al. 1993). Humans have a long history of modification of aquatic habitats through alteration of wetlands, addition of agricultural runoff and small to large-scale fishing activities, which in turn may influence habitat quality, fish abundance and diversity of waterbird assemblages (Fleury and Sherry 1995; Klein et al. 2008; Atkinson et al. 2010; Lunardi and Macedo 2013).
Fishery activities continue to contribute significantly towards biodiversity loss (Klein et al. 2008) although noteworthy ways of sustainable use of fish resources, such as utilization of ecosystem-based management approaches, are currently being explored as options (Kellner et al. 2011; Hamel et al. 2013). There is evidence showing that attaining sustainability in fisheries is possible through good governance involving ecosystem-based management approaches (Green and Elmberg 2014; Bundy et al. 2017). However, there is a major gap in our understanding of how small-scale, traditional fishing practices influence biodiversity (Oliveira Júnior et al. 2016). Small-scale fisheries employ a wide variety of gear types including traps, gillnets, and drift gillnets, which vary in the way they interact with waterbirds (Morgan and Chuenpagdee 2003; Pravin et al. 2011). The type of gear could be advantageous for the hunting strategies of certain waterbirds, for example, while detrimental to others (e.g. De Nie 1995; Battley et al. 2003; Lunardi and Macedo 2013). Comparison of the impact of the types of fishing gear may help to assess negative impacts on certain species (e.g. bycatch of pursuit diving waterbirds) or positive impacts on piscivorous littoral waterbirds (e.g. herons and egrets through facilitation of fishing opportunities). Assessments of gear type could help in selecting gear types that minimize detrimental effects on aquatic species assemblages (Crivelli 1990), consistent global efforts to promote sustainable use of biodiversity and reduction of biodiversity loss (CBD 2010).
Fishing is a major source of income and protein for millions of people in the Indian subcontinent, with the vast majority engaged in some forms of small-scale inland fishery (Pravin et al. 2011). A wide range of artisanal or traditional fishing gears are available and are currently in use in inland fisheries throughout the subcontinent, including bamboo traps, gill nets, and seines (Pravin et al. 2011). Bamboo traps, one of the oldest types of gear used in the Indian subcontinent, are mainly operated in rivers and backwaters with more focus in the middle and upper stretches of the river where the operation of other fishing gears like gill nets and seines are difficult. Bamboo traps are passive fishing gear and are designed to permit entry of fish but prevent their exit, causing entrapment (Pravin et al. 2011). Bamboo traps are operated continuously with periodic checking to remove entrapped fish that remain alive without injury. Although traditional fishing practices are often considered better for biodiversity conservation, the extent and scale of traditional fishing may still incur negative consequences on biodiversity, especially since increasing population size in developing countries imposes a cumulative impact on fish resources with cascading effects on aquatic ecosystems (Tittensor et al. 2014; Oliveira Júnior et al. 2016; Bundy et al. 2017). An understanding of the impacts of traditional fishing practices on other biodiversity is limited throughout the developing countries and is an important research gap that needs to be addressed to achieve sustainability in fisheries and biodiversity conservation (Oliveira Júnior et al. 2016; Bundy et al. 2017).
The aim of this study was to evaluate the impact of two traditionally used fishing methods on the waterbird assemblages in a globally important wetland area in south western India and to determine if such traditional fishing practices could reduce pressures on and even improve the state of biodiversity.
The Kadalundi-Vallikkunnu Community Reserve (KVCR, 11°7′28″-11°8′01″N, 75°49′36″-75°50′20″E) is located at the mouth of the River Kadalundi that drains into the Arabian Sea on the west coast of Kerala (Fig. 1). The river divides into two channels encircling a small island before entering the sea (Uthaman and Namasivayan 1991). Apart from scattered patches of mangroves, the estuary is bordered by coconut groves and human habitation. Around eight hectares of mudflat—exposed during low tides—offer potential foraging ground for several hundreds of wintering and resident waterbirds, particularly shorebirds (Aarif et al. 2011, 2015). The Kadalundi estuary is an important stopover and wintering ground in the west coast of India. The area was declared as the Kadalundi-Vallikkunnu Community Reserve (KVCR) in 2007 due to its importance (Aarif and Prasadan 2014). Earlier studies showed that this site has undergone significant declines in shorebird assemblages although shorebirds seem to have adapted to changing patterns in habitat type by altering habitat use patterns (Aarif et al. 2014). The area provides significant socio-economic and livelihood services (fishing, oyster farming and sand mining) for the people living around the estuary. Bamboo traps are the most commonly used traps in the Kadalundi estuary. Various types of fishing methods are operated in the Kadalundi estuary for commercial exploitation of the fishery resources and there is evidence showing that waterbirds are attracted to fishing gear causing increase in numbers and diversity of certain waterbird assemblages.
Three major habitats could be distinguished in the KVCR wetland, namely mudflats, mangroves and sand beds. Mudflats are fully exposed during low tide. The mangrove forests are located in the eastern side of the mudflat. Sand beds are extensions of mudflats but are entirely composed of sandy sediment.
The present study was conducted from January 2012 to December 2015. Four visits were made every month and direct waterbird counts were done using a binocular (10 × 50) during low tide following Hoves and Bakewell (1989). Of the four monthly visits, the first and third visits were made during which fishermen had their traps deployed whereas the second and fourth visits were during periods without any trapping activities. The present study included the following common species in the area: Little Egret (Egretta garzetta), Intermediate Egret (Mesophoyx intermedia), Great Egret (Casmerodius albus), Western Reef Egret (Egretta gularis), Indian Pond Heron (Ardeola grayii), Grey Heron (Ardea cinerea), Little Green Heron (Butorides striatus), Little Cormorant (Phalacrocorax niger), Darter (Anhinga melanogaster), Small Blue Kingfisher (Alcedo atthis), Stork-billed Kingfisher (Halcyon capensis), White-breasted Kingfisher (Halcyon smyrnensis), Oriental White Ibis (Threskiornis melanocephalus), White Breasted Waterhen (Amaurornis phoenicurus), and Brahminy Kite (Haliastur indus). Total counts of these species were made in both periods when nets and bamboo cages were either deployed or not deployed.
The nylon nets ranged in length from 100 to 1000 m and with a width of 1.5 m, which are fixed on bamboo stakes. The nets are deployed on the estuary ten times every month at mid night. Cages made of bamboo ropes, are also deployed along side the nets. Large fishes are collected following morning at low tide from the both ends of the net and from the bamboo cages. Even during the low tide sufficient water remains in the cages to allow the fishes to remain alive. The cages are covered with polythene nets to prevent the waterbirds from catching the trapped fish. Waterbirds commonly wait on top of the nets to opportunistically capture trapped fish. Fish of all sized are trapped in the cages although the fishermen only collect the larger fishes while the smaller fishes are released back into the estuary. Fish collection from the nets takes about 1-3 h after which the nets are removed slowly. During this period, the waterbirds congregate near the nets to feed upon the small fishes which are discarded by the fishermen.
Total fish collected by all the fishermen was weighed at the end of fish collection two times a week, corresponding with a trap deployment period. We used quantity of fish captured as an index of fish population size (Pope et al. 2010), fully recognizing some of the concerns of linking fish populations to catch. We used this index to determine temporal changes in fish populations as well as their linkage with waterbird abundance and diversity.
The count data were summarized and categorized by years (2012-2015), seasons (pre-monsoon, monsoon, post-monsoon) and period of fishing traps (traps deployed, traps not deployed). We used multi-way analysis of variance (factorial ANOVA) to examine how waterbird counts and waterbird diversity (measured as Shannon-Weiner index of diversity, H) varied in relation to years, seasons and period of fishing traps. In each case, we checked for interactions in the dependent variables (year, season and period). We also used one-way ANOVAs to test for differences in the number of waterbirds between selected habitats (mudflats, mangroves, sand bed) and to highlight the variation of the quantity of captured fish (Kg per month for each study year) between years. Finally, in order to examine the relationship between fish occurrence and waterbird counts and diversity, we conducted a linear regression with waterbird count or waterbird diversity as the response variables and quantity of fish as explanatory variable.
Waterbird counts varied significantly between years (F = 12.53, df = 3, p < 0.001), seasons (F = 48.40, df = 2, p < 0.001) and particularly with period of fishing activities (trap deployment) (F = 94.73, df = 1, p < 0.001) (Table 1). Significant interactions were observed between years and period and seasons and period but not between years and seasons (Table 1). Thus, we interpreted the significant relationships between bird counts and the three explanatory variables based on the interaction terms in the model (Sokal and Rohlf 2012). The largest waterbird count was recorded in 2012. It also decreased from post-monsoon through pre-monsoon to monsoon (Fig. 2). The highest values of waterbird counts were recorded during the trap deployment period (Fig. 3).
| Dependent variable | df | F | p |
| Bird counts | |||
| Intercept | 1 | 269.70 | < 0.001 |
| Years | 3 | 12.53 | < 0.001 |
| Seasons | 2 | 48.40 | < 0.001 |
| Period | 1 | 94.73 | < 0.001 |
| Years * seasons | 6 | 1.08 | 0.37 |
| Years * period | 3 | 3.96 | < 0.01 |
| Seasons * period | 2 | 17.76 | < 0.001 |
| Years * seasons * period | 6 | 0.12 | 0.99 |
| Error | 168 | ||
| Bird diversity | |||
| Intercept | 1 | 1448.43 | < 0.001 |
| Years | 3 | 2.31 | 0.07 |
| Season | 2 | 33.43 | < 0.001 |
| Period | 1 | 100.87 | < 0.001 |
| Years * season | 6 | 0.82 | 0.55 |
| Years * period | 3 | 0.50 | 0.67 |
| Season * period | 2 | 1.09 | 0.33 |
| Years * season * period | 6 | 0.90 | 0.49 |
| Error | 168 | ||
 DownLoad:
CSV
DownLoad:
CSV
The waterbird diversity was significantly higher during the trap deployed period (F = 100.87, df = 1, p < 0.001) (Table 1, Fig. 3). The highest and the lowest values of diversity index (Shannon index) were recorded in post-monsoon and monsoon, respectively (F = 33.43, df = 2, p < 0.001) (Fig. 4). However, no variation of waterbird diversity was found between years (F = 2.31, df = 3, p = 0.07) and there were no interactions between years, seasons and periods (Table 1).
The highest waterbird count was recorded in the mudflats. It was significantly larger than those noted at mangroves and sand beds (one-way ANOVAs, F = 106.95, df = 2, p < 0.001) (Fig. 5). Although the waterbird diversity measured at the mudflats appeared higher than that recorded at others habitats, this variation was not significant (F = 0.24, df = 2, p = 0.78) (Fig. 5). Individual species varied in their responses to trap deployment. The Oriental White Ibis (one-way ANOVAs, F = 7.73, p < 0.001), the Little Cormorant, the Little Green Heron, the Little Egret, the Intermediate Egret, the Indian Pond Heron and the Western Reef Egret increased in abundance when traps were deployed (one-way ANOVAs, p < 0.05, in all cases). The Great Egret, Grey Heron and Small Blue Kingfisher did not change in abundance in relation to trap deployment (one-way ANOVAs, p > 0.05, in all cases). The Brahminy Kite increased in abundance when traps were not deployed (one-way ANOVA, F = 7.96, p < 0.01).
The quantity of captured fish significantly decreased from 2012 to 2015 (one-way ANOVAs, F = 10.61, df = 3, p < 0.001) (Fig. 6). Furthermore, the waterbird counts (Linear regression, F = 38.83, df = 1, p < 0.001) and the diversity (Linear regression, F = 16.51, df = 1, p < 0.001) were significantly increased based on fish captured.
Achieving sustainability in fisheries and safeguarding associated biota in aquatic ecosystems are major goals in conservation science (CBD 2010; Oliveira Júnior et al. 2016; Bundy et al. 2017). Small-scale, traditional fishing activities are carried out worldwide and their impact on aquatic ecosystems seems to be high, given increasing populations and increasing fishing effort (Hamel et al. 2013). Fishing or fishery activities are often associated with enhanced waterbird abundance and diversity (Atkinson et al. 2010; Lunardi and Macedo 2013). Fishing traps or social foraging activities may restrict prey fish species to confined, shallow areas, serving as attractants for waterbirds and increasing success of predation (Battley et al. 2003). In this study, we showed that waterbird abundance and diversity at KVCR were positively influenced by traditional fishing activities.
The distribution pattern of waterbirds is strongly influenced by habitat conditions and availability of food in wetlands (Goss-Custard et al. 1977; Fonseca et al. 2004; Green and Elmberg 2014). Shorebirds are specific in their choice of habitats and spatial distribution varies with availability of food resources (Frederickshon and Reid 1990; Skagen and Knopf 1993; Pandiyan and Asokan 2015). In altered or impoverished habitats waterbirds supplement their energy intake by using multiple adjacent wetlands within the landscape (Dunning et al. 1992). In this study, waterbird counts varied significantly between years and seasons due to both natural and anthropogenic factors. Highest waterbird counts were recorded in 2012 and especially during the post-monsoon periods. The number of waterbirds decreased successively from post-monsoon to monsoon seasons. A high seasonality in the waterbird composition and abundance was observed that coincided with seasonal variations in resource availability (Mundava et al. 2012). The natural dynamics of waterbird populations are mainly driven by rainfall in much of South and Southeast Asia (Gopal and Chauhan 2001; Aarif et al. 2014). The southwest monsoons in the Indian Subcontinent deposits substantial amounts of rain causing an expansion in the spatial area of rivers, lakes, reservoirs and wetlands. Waterbirds are presented with an abundance of food over a vastly enhanced aquatic resource causing them to disperse over a wide area. Thus, they do not congregate in small water bodies, causing a decline in their counts. Additionally, we recorded the highest waterbird counts in mudflats compared to the mangroves and sand beds in KVCR. This is also consistent with earlier studies showing switches in habitat use patterns in shorebirds due to apparent changes in the quality and extent of these habitats (Aarif et al. 2014).
We were able to demonstrate an increase in waterbird abundance and diversity relative to trap deployment consistently between years, seasons and habitat types. The traditional fishing practice of laying out nets and bamboo traps had an overall positive effect on waterbird assemblages. Fish discarded by fishermen were small-sized (3-5 cm long, particularly Grey Mullet, Mugil cephalus; Aarif, personal observation) and this attracted large numbers of the common waterbirds to these areas during trap deployment periods that were taking advantage of shallow areas where fish were trapped. Several studies show that congregation of waterbirds may occur in response to seasonal increase in the abundance of fish (De Nie 1995; Fleury and Sherry 1995; Warke and Day 1995; Battley et al. 2003). Cormorants of many species increase drastically in population size in response to increased fish stocks (De Nie 1995; Warke and Day 1995). Social foraging of mixed species assemblages of herons, egrets and long-legged shorebirds concentrate fish into small, shallow areas allowing active predation (Battley et al. 2003). Fleury and Sherry (1995) showed that crayfish fisheries increased the abundance of a variety of egrets and herons, including the Great Blue Heron, Great Egret, Black-crowned Night Heron, and several others, that used crayfish as a major part of their diet. In another study by Lunardi and Macedo (2013), manual collection of shellfish in coastal Australia was followed by increased foraging by shorebirds, suggesting that disturbance caused by humans improved predation. We found that the Oriental White Ibis, Little Cormorant, Little Green Heron, Little Egret, Intermediate Egret, Indian Pond Heron and Western Reef Egret (relatively small with length ranging from 370 to 500 mm) increased during trap deployment, because these species typically hunt in mixed flocks in shallow shorelines. The larger birds like the Grey Heron, Great Egret (810‒995 mm total length), and the Small Blue Kingfisher were not affected due to trap deployment. This could be because these species prefer slightly deeper, more open waters for hunting, although they may forage in mixed flocks of other smaller herons and egrets, including the species above (Rasmussen and Anderton 2005). Similarly, the Small Blue Kingfisher hunts by diving into the water from a perch, a method that may not be benefitted by the deployment of traps, especially those that are already occupied by many other waterbirds. Thus, the small waterbirds with hunting strategies involving shallow water were benefitted by the deployed traps. What impact would the enhancement of some species have on the community structure in this aquatic food web? This is a question that needs to be addressed more carefully with further study. Enhancement of some of species could mark the path towards sustainable fishing practices, where fish harvests were controlled and avian biodiversity was not experiencing significant and long-term declines.
Peak season for bamboo fish trap deployment was observed during December to May (Remesan and Ramachandran 2008) which coincides with the peak number of waterbirds in the post-monsoon season in the KVCR. Waterbird abundance and diversity were significantly related to fish capture, providing further support to the fact that fishing activities in the post-monsoon period could enhance waterbird assemblages even further. Small fishes like mullets, small catfish, prawns and other medium and small-sized fishes which are favored by waterbirds like egrets and herons congregate near the edges of the traps. On the other hand, juveniles of many fishes constitute the major share of the bycatch (Remesan and Ramachandran 2008) and we observed 17 species of fishes (data not shown) of which the dominant groups include thirutha/kanambu (Liza sp./Mugil spp.), chemballi (Lutjanus sp.), koori/etta (Mystus sp./Arius sp.), pearl spot (Etroplus spp.), grey mullet (M. cephalus), and prachi (Gerres spp.). Thus, a wide range of fish become available to forage by waterbirds near the traps causing them to flock in large numbers during trap deployment, especially during the post-monsoon period when trap deployment is intense. This is consistent with other studies, for example, total crayfish landings in the Louisiana crayfish industry was related to increases in abundance and diversity of egrets, herons and other waterbirds (Fleury and Sherry 1995). The quantity of captured fish significantly decreased from 2012 to 2015 probably due to overfishing, insufficient management and habitat degradation, pollution (Bijoy 2008) and nutrient enrichment (Aarif et al. 2014). Providing high quality habitats for waterbirds as well as effective sustainable livelihood through traditional fisheries is of critical importance to biodiversity conservation (Weber and Haig 1996; Erwin 2002; Taft et al. 2002; Klein et al. 2008; CBD 2010; Mcowen et al. 2016). The interactions between waterbirds and traditional fish traps have not caused any mortality of waterbirds in the KVCR. The variation in habitat requirements among waterbird species and groups suggests that wetland management must be based on region-specific knowledge about waterbird communities. The managers also need to understand the seasonal dynamics of waterbirds, so that management can be timed to meet the special needs of breeding, stopover, and wintering periods (Isola et al. 2002; Parsons 2002). Kadalundi estuary remains an important stopover for migrating waterbirds that is under constant threat from anthropogenic sources. We present evidence that trapping methods could work synergistically to support local avifauna. Proper management of the fish resources through ecosystem based management practices, utilizing traditional fishing methods, and recognizing and safeguarding important areas supporting migratory shorebirds could help to achieve longterm sustainability of the fisheries and the biodiversity of which they are a part.
In this study we documented a positive association between traditional fish trap deployment and waterbird counts and diversity. This suggests that the traditional fishing practices in KVCR, if promoted and managed adequately, could enhance waterbird abundance and diversity. Low impact fishing practices such as the bamboo fish traps that enhance waterbird abundance could serve as a model for sustainable utilization of fish resources, provided proper monitoring and evaluation instruments are in place (Tittensor et al. 2014; Mcowen et al. 2016).
KMA and SBM conceived of the study and designed the experiments. KMA, KKM and PKP conducted the experiments in the field. AN and KMA carried out the analysis part. KMA, PKP, AN and KKM wrote the first draft of the manuscript. SBM supervised the research and revised the draft. All authors read and approved the final manuscript.
The authors declare that they have no competing interests.
|
Fritz J. Reintroduction of the Northern Bald Ibis in Europe: Illegal hunting in Italy during autumn migration as the main threat. WAZA Mag. 2015; 31.
|
|
Fritz J, Unsoeld M, Voelkl B. Back into European wildlife: the reintroduction of the Northern Bald Ibis (Geronticus eremita). In: Kaufman A, Bashaw M, Maple T, editors. Scientific foundations of zoos and aquariums: their role in conservation and research. Cambridge: Cambridge University Press; 2019. p. 339–66.
|
|
Fritz J, Unsöld M. Internationaler artenschutz im kontext der IUCN reintroduction guidelines: Argumente zur wiederansiedlung des waldrapps Geronticus eremita in Europa. Vogelwarte. 2015; 53: 157–68.
|
|
Nagata H, Yamagish S. Re-introduction of crested ibis on Sado Island, Japan. In: Soorae PS, editor. Global re-introduction perspectives: 2013. Further case studies from around the globe. Gland, Switzerland: IUCN/SSC Re-introduction Specialist Group and Abu Dhabi, UAE: Environment Agency-Abu Dhabi; 2013. p. 58–62.
|
|
Pschera A. Das Internet der Tiere. Berlin: Matthes & Seitz Verlag; 2014.
|
|
Tahvanainen K, Niño J, Halonen P, Kuusela T, Alanko T, Laitinen T, et al. Effects of cellular phone use on ear canal temperature measured by NTC thermistors. Clin Physiol Funct. 2007; I(27): 162–72.
|
| 1. | Prateek, Mishra, H., Kumar, V., Kumar, A. Population structure, distribution pattern and habitat utilization of Yellow-wattled Lapwing, Vanellus malabaricus (Boddaert, 1783), in Northern India. BMC Zoology, 2025, 10(1): 1. DOI:10.1186/s40850-025-00222-6 |
| 2. | Aarif, K.M., Nefla, A., Rubeena, K.A. et al. Assessing environmental change and population declines of large wading birds in southwestern India. Environmental and Sustainability Indicators, 2025. DOI:10.1016/j.indic.2024.100572 |
| 3. | Barman, A., Rajak, F., Jha, R. Integrating Traditional Knowledge Systems for Wetland Conservation and Management: A Critical Analysis. Nature Environment and Pollution Technology, 2025, 24(1): B4212. DOI:10.46488/NEPT.2025.v24i01.B4212 |
| 4. | Rubeena, K.A., Aarif, K.M., Bjedov, D. et al. Species-Specific Accumulation and Temporal Variation of Metal(loid)s Shape the Population Trends of Large Wading Birds in the Eastern Province of Saudi Arabia. Archives of Environmental Contamination and Toxicology, 2025. DOI:10.1007/s00244-025-01162-5 |
| 5. | Himanshu Mishra, Bano, F., Prateek, Kumar, A., Kumar, V. Elucidation of Diversity and Habitat Utilization of Waterbirds in Khajuha Wetland, Northern India. Biology Bulletin, 2024, 51(3): 800-812. DOI:10.1134/S1062359023605542 |
| 6. | Rutina, L.P., Simasiku, E., Kabanze, J.M. Piscivore water bird diversity at freshwater tributaries of Zambezi River, Namibia. Koedoe, 2024, 66(1): a1815. DOI:10.4102/koedoe.v66i1.1815 |
| 7. | Aarif, K.M., Rubeena, K.A., Nefla, A. et al. Heavy metals in wetlands of southwestern India: from sediments through invertebrates to migratory shorebirds. Chemosphere, 2023. DOI:10.1016/j.chemosphere.2023.140445 |
| 8. | Anand, J., Byju, H., Nefla, A. et al. Conservation significance of Changaram wetlands - a key wintering site for migratory shorebirds and other waterbirds in the western coast of Kerala, India. Journal of Threatened Taxa, 2023, 15(1): 22410-22418. DOI:10.11609/jott.8089.15.1.22410-22418 |
| 9. | Rubeena, K.A., Nefla, A., Aarif, K.M. et al. Alterations in hydrological variables and substrate qualities and its impacts on a critical conservation reserve in the southwest coast of India. Marine Pollution Bulletin, 2023. DOI:10.1016/j.marpolbul.2022.114463 |
| 10. | Athira, T.R., Nefla, A., Shifa, C.T. et al. The impact of long-term environmental change on zooplankton along the southwestern coast of India. Environmental Monitoring and Assessment, 2022, 194(4): 316. DOI:10.1007/s10661-022-09921-w |
| 11. | Aarif, K.M., Nefla, A., Nasser, M. et al. Multiple environmental factors and prey depletion determine declines in abundance and timing of departure in migratory shorebirds in the west coast of India. Global Ecology and Conservation, 2021. DOI:10.1016/j.gecco.2021.e01518 |
| 12. | Franco-Moraes, J., Clement, C.R., Cabral de Oliveira, J. et al. A framework for identifying and integrating sociocultural and environmental elements of indigenous peoples’ and local communities’ landscape transformations. Perspectives in Ecology and Conservation, 2021, 19(2): 143-152. DOI:10.1016/j.pecon.2021.02.008 |
| 13. | Zhang, P., Li, Y., Tian, W. et al. STUDY ON THE DYNAMIC CHANGE OF WATERBIRD DIVERSITY AND DISTRIBUTION IN XIANGHAI. International Geoscience and Remote Sensing Symposium IGARSS, 2021. DOI:10.1109/IGARSS47720.2021.9554821 |
| 14. | Ciobota, M., Murariu, D., Ciobota, A. Traditional Fish Pond Management Provides Reliable Foraging Opportunities for Migrating Black Storks in Romania. Ardea, 2020, 108(2): 191-201. DOI:10.5253/arde.v108i2.a10 |
Figures(3) / Tables(2)





| Dependent variable | df | F | p |
| Bird counts | |||
| Intercept | 1 | 269.70 | < 0.001 |
| Years | 3 | 12.53 | < 0.001 |
| Seasons | 2 | 48.40 | < 0.001 |
| Period | 1 | 94.73 | < 0.001 |
| Years * seasons | 6 | 1.08 | 0.37 |
| Years * period | 3 | 3.96 | < 0.01 |
| Seasons * period | 2 | 17.76 | < 0.001 |
| Years * seasons * period | 6 | 0.12 | 0.99 |
| Error | 168 | ||
| Bird diversity | |||
| Intercept | 1 | 1448.43 | < 0.001 |
| Years | 3 | 2.31 | 0.07 |
| Season | 2 | 33.43 | < 0.001 |
| Period | 1 | 100.87 | < 0.001 |
| Years * season | 6 | 0.82 | 0.55 |
| Years * period | 3 | 0.50 | 0.67 |
| Season * period | 2 | 1.09 | 0.33 |
| Years * season * period | 6 | 0.90 | 0.49 |
| Error | 168 | ||
DownLoad:
CSV





 Email Alerts
Email Alerts RSS Feeds
RSS Feeds