Fengting ZHAO, Lizhi ZHOU, Wenbin XU. 2013: Habitat utilization and resource partitioning of wintering Hooded Cranes and three goose species at Shengjin Lake. Avian Research, 4(4): 281-290. DOI: 10.5122/cbirds.2013.0032
Citation:
Fengting ZHAO, Lizhi ZHOU, Wenbin XU. 2013: Habitat utilization and resource partitioning of wintering Hooded Cranes and three goose species at Shengjin Lake. Avian Research, 4(4): 281-290. DOI: 10.5122/cbirds.2013.0032
Fengting ZHAO, Lizhi ZHOU, Wenbin XU. 2013: Habitat utilization and resource partitioning of wintering Hooded Cranes and three goose species at Shengjin Lake. Avian Research, 4(4): 281-290. DOI: 10.5122/cbirds.2013.0032
Citation:
Fengting ZHAO, Lizhi ZHOU, Wenbin XU. 2013: Habitat utilization and resource partitioning of wintering Hooded Cranes and three goose species at Shengjin Lake. Avian Research, 4(4): 281-290. DOI: 10.5122/cbirds.2013.0032
The loss and degradation of wetlands have adversely affected waterbirds, which depend on wetland habitats. Interspecific competition has an important effect on habitat utilization of wintering waterbirds. Resource utilization, including partitioning, in degraded wetlands has become a hot issue in ecological studies of wintering waterbirds. In order to have an insight into the habitat utilization and resource partitioning between a Hooded Crane (Grus monacha) population and the guild of three goose species, i.e., Anser fabalis, A. albifrons and A. erythropus wintering in lake wetlands, we carried out a study at Shengjin Lake National Nature Reserve from November 2011 to April 2012. We surveyed the Hooded Cranes and goose guild foraging in various habitats during the wintering periods with a combined method of fixed route searching and fixed site observations. Resource partitioning was studied by means of calculating habitat utilization rates and the width and overlap of spatial niches. The results showed that the habitat utilization rate and the width of spatial niches of the Hooded Crane population and goose guild shifted with the season. The habitat utilization rates of the cranes in grasslands were high at all three wintering stages. The habitat utilization rates were 0.454, 0.435 and 0.959 respectively for the Hooded Cranes and 0.627, 0.491 and 0.616 for the goose guild. This suggests that the overlap in grasslands was higher between cranes and goose guild. Most habitats were accessible at the middle stage, so the width of the spatial niche of the cranes (1.099) and goose guild (1.133) both reached their peak at this stage. The greatest niche overlap was 0.914 for these two groups at the late stage, followed by 0.906 at the middle stage and the smallest was 0.854 at the early stage. Ecological response to the changes in habitats of wintering waterbirds was clearly shown in the dynamic variations of the niche of both the Hooded Cranes and the three goose species. Coexistence among waterbirds was achieved by regulation of niche width to reduce niche overlap and relieve interspecific resource partitioning.
The lake wetlands in the middle and lower Yangtze River floodplain are important stopover and wintering sites for migratory waterbirds on the East Asian-Australian flyway. In recent years, due to the over-exploitation of lake resources, wetland degradation has become a serious issue and habitats, suitable for these migratory waterbirds, are disappearing. Food resource utilization is characterized by interspecific competition among wintering waterbirds, which may be intensified by food shortages (Xiang and Wang, 2005; Jiang et al., 2007; Wang et al., 2011). The wintering period is a critical period in the yearly cycle of waterbirds. Sufficient energy supplements in this period are critical to their migration, reproduction and survival (Morrison et al., 2007; Anna, 2010; Both et al., 2010). Food, space and other resources are extremely limited in degraded wetlands, especially in cold and severe winters. Their competitive intensity usually reaches its peak at this time (Oksanen, 1987). The mechanism of resource partitioning for waterbirds inhabiting the lake wetlands has attracted considerable attention of late.
Species coexist through resource partitioning, including the partition of habitat, food and habitat utilization time (Schoener, 1974; Mittelach, 1984; Reinert, 1984). Variations of habitat utilization and foraging behavior of coexisting species have been considered evolutionary strategies to partition limited resources and to minimize potential interspecific competition (Jenni, 1969; Willard, 1977; Ishtiaq, 2010). Temporal-spatial variations of habitats are the main factors affecting habitat utilization (Kelly et al., 1996; Ribeiro, 2004). In a highly degraded wetland ecosystem, common resources are shared by large flocks of migratory waterbirds. These high-density waterbird flocks may induce greater interspecific competition (Burger et al., 1977; Alatalo et al., 1985; Beerens et al., 2011). As a result, significant differences may be found in microhabitat utilization and food selection (Davis and Smith, 2001; Vahl et al., 2005; Kober and Bairlein, 2009).
Cranes and geese are common wintering waterbirds inhabiting the lakes of the middle and lower Yangtze River floodplain. The Hooded Crane (Grus monacha) is a vulnerable species (VU) on the IUCN red list and a species of wildlife under first class state protection in China. The Bean Goose (Anser fabalis), the Greater White-fronted Goose (A. albifrons) and the Lesser White-fronted Goose (A. erythropus) are three common goose species in the wetlands of the middle and lower Yangtze River floodplain. They are regarded as indicator species of lake ecosystems due to their sensitivity to habitat change. These waterbirds migrate southwards to the wintering grounds in the middle and lower reaches of the Yangtze River in late October every year. The main habitat of Hooded Cranes and these three goose species in winter are the grasslands, farmlands and shallow-water areas. In general, the wintering Hooded Cranes occupy the dry lakeshores, grasslands and paddy fields. They mainly feed on tubers of Vallisneria natans, as well as on seedlings of Polygonum lapathifolium, occasionally on Potamogeton malainus, Phalaris arundinacea, Cynodom dactylon, Carex unisexualis, Cyperus sp., as well as on wheat seedlings, rice grains, spiral shells and mussels. Their diet is affected by the availability of food resources in their wintering habitat (Wang and Hu, 1986; Liu et al., 2001). The three goose species often assemble in mixed flocks, as a guild with extremely similar characteristics of habitat utilization (Yang, 2011; Chen and Zhou, 2011). Some of their food resources overlap with those of the Hooded Cranes, such as Carex spp., Phalaris arundinacea, rice grains and Cynodomdactylon (Fox et al., 2008; Zhao et al., 2010). Hence, geese are the major competitors of Hooded Cranes. This study aims: (a) to gain an insight into the characteristics of habitat utilization of wintering Hooded Cranes and the goose guild in lake wetlands and (b) to explore resource partitioning between Hooded Cranes and the goose guild.
Methods
Study area
Shengjin Lake (116°55′–117°15′E, 30°15′–30°30′N), located to the south of the Yangtze River bank, is a shallow-water, river-connecting lake. A river is connected to the lake via the Huangpen Sluice built in 1965. The lake area is at its largest at 14000 ha in the high water season, while it is smallest at 3400 ha in the low water season. The lake usually is divided into two parts: the upper part and lower part from south to north. The region where the lake is located belongs to the northern subtropical humid zone with an annual mean temperature of 16.1 ℃ and precipitation of 1600 mm. Shengjin Lake is one of the most important wintering and stopover grounds along the Yangtze River for waterbirds, especially Hooded Cranes, Bean Geese, Greater White-fronted Geese and Lesser White-fronted Geese. As one of the major wintering grounds for migratory waterbirds, it was approved in 1986 to be established as a provincial nature reserve and in 1997 it became a national nature reserve.
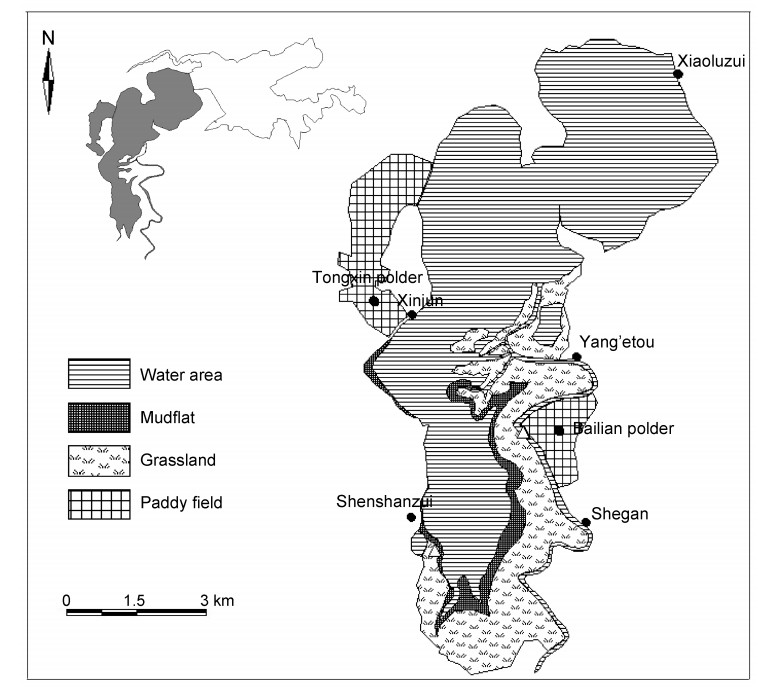
The present study was carried out in the upper part of Shengjin Lake, which is located in the southern part, a core area of this nature reserve. The lake bed is smooth and flat, while the terrain is higher towards the southeast. Cage aquaculture operations had been established at Shengjin Lake for more than ten years, while pond and enclosure culture are commonly seen in the lake. In the low water season, the lake water retreats and a large mudflat is exposed to provide a foraging habitat for the Hooded Crane and other wintering waterbirds. The foraging habitats, consisting of water areas, mudflats, grasslands and paddy fields (Fig. 1), show periodic and dynamic changes.
Figure
1.
Foraging habitats for Hooded Cranes and other waterbirds at the upper part of Shengjin Lake
The habitat utilization of Hooded Cranes and geese are affected by periodic changes in the hydrology and mudflat exposure of Shengjin Lake. We divided the wintering period into three stages according to the hydrological variations in the lake. The early stage was before late December. During this time the lake, still at a high water table, started to recede. The middle stage was from the early January to late February in the following year, when the water level dropped quickly and large areas of the lake shore became exposed. The late stage was from late February to the end of March, during which the lake shore had become dry and the water level began to rise again.
The habitat types of the upper Shengjin Lake varied with the hydrological conditions during the low water season. At the middle stage, habitats were plentiful, including deep-water areas, shallow-water areas, mudflats, grasslands and paddy fields. The deep-water area in the lake was only 0.5 m deep. The shallow-water area refers to the water body with a depth of less than 0.5 m. The mudflats are tidal flats with large areas of the lake beach exposed, when the water has retreated. Some smartweeds, sedges and other plants grow in areas where the water retreats early. Grasslands are mudflats with a vegetation cover of more than 20%, dominated by herbaceous plants. Paddy fields are in polders where rice is planted in the spring and summer and harvested in autumn and then wheat is planted in late winter.
Foraging habitats for Hooded Cranes and geese are relatively stable at a certain wintering stage at Shengjin Lake (Cao et al., 2010; Zhou et al., 2010). These foraging habitats are centralized in Xinjun village, as well as in the range between Yang'etou and Shegan villages (Fig. 1).
Data collection
We combined the methods of fixed route and fixed site observations to investigate the habitats at the upper part of Shengjin Lake from November 15, 2011 to April 1, 2012. Each survey covered the whole array of habitat types. Four routes were included in one sampling survey, which lasted from 1 to 3 days. Sampling surveys which were not completed were excluded from the final samples. Therefore, the sample sizes of Hooded Crane and geese were equal. The valid data collection consisted of a total of 59 days and 38 samples, including 15 samples collected in 30 days for the early wintering stage, 13 samples collected in 19 days for the middle wintering stage and 10 samples in 10 days for the late wintering stage. The distribution of Hooded Cranes and geese at the upper lake were determined by means of a fixed route survey. The surveyed routes were from Xinjun to Xiaoluzui village, Xiaoluzui to Shegan village, Xinjun to Shenshanzui village and from Shenshanzui to Shegan village. Fixed site observatione were carried out when we focused on Hooded Cranes and/or geese. The area within a radius of 1 km was observed by a binocular telescope (BOWAS 8 × 42) and a monocular telescope (SWAROVSKI 20-60 × 80); we recorded the number and habitat types. A direct counting method was employed for small number of cranes and geese (generally less than 300 birds). For large flocks, a group counting method was adopted. That is, the flock was divided into several smaller individual groups such as 10, 50 to 100. The number of birds in the whole flock was estimated by counting the number of birds in each smaller flock (Howes and Bakewill, 1989; Ma, 2006). Since the diets of the three goose species are similar and often mixed in high-density flocks, the three goose species were treated as one guild for the purpose of counting.
Data analysis
Based on the distribution of the populations of Hooded Cranes and geese in each survey, the utilization rates (U) of all habitat types by Hooded Crane or goose guild were calculated as: Ui = Ni/N, where Ui is the utilization rate of the ith habitat type by waterbirds; Ni the number of the waterbirds in the ith habitat type and N the total number of waterbirds in all habitat types.
The mean utilization rate (Mean) and standard error (SE) in all habitats and all wintering stages were calculated.
The utilization rates of habitats by the waterbirds at the same wintering stage were compared with a Kolmogorov-Smirnov test in SPSS17.0. The utilization rates in five habitat types were checked to see if the assumption of a normal distribution were met. If met (p > 0.05), an independent-sample t test was performed; if not (p < 0.05), the Mann-Whitney U test was used. The significance level was set as α = 0.05.
where Bi is width of the niche; Pi the percentage of individual birds observed in the ith habitat type from the total number of Hooded Cranes or goose guild.
where Pjk and Pjk are the proportions of Hooded Crane (i) and goose guild (j) observed in the kth habitat type.
Oij=0whennonicheoverlap;Oij=1whencompleteoverlap.
Results
Habitat utilization of Hooded Crane population and goose guild
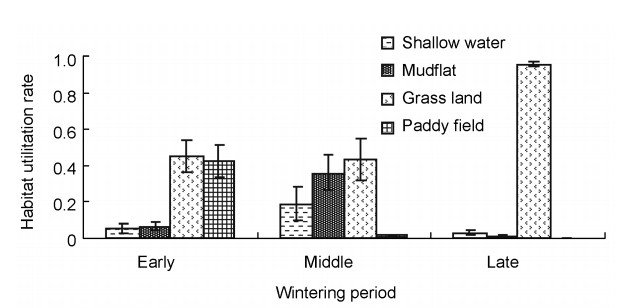
The four major habitat types for Hooded Cranes at Shengjin Lake consisted of shallow-water areas, mudflats, grasslands and paddy fields. The most utilized habitat type was grasslands at the early wintering stage, with a utilization rate of 0.454 ± 0.083 (n = 15), followed by 0.427 ± 0.088 (n = 15) for the paddy fields. The utilization rates of shallow-water areas and mudflats were relatively low, i.e., 0.053 ± 0.024 (n = 15) and 0.066 ± 0.021 (n = 15), respectively. The most frequently utilized habitats were grasslands with a utilization rate of 0.435 ± 0.115 (n = 13) and mudflats of 0.363 ± 0.101 (n = 13) at the middle wintering stage, followed by 0.190 ± 0.091 (n = 13) for the shallow-water areas and 0.012 ± 0.008 (n = 13) for paddy fields. The utilization rate of grasslands was 0.959 ± 0.015 (n = 10), which was clearly higher than that of other habitats at the late wintering stage. The utilization rate was 0.033 ± 0.011 (n = 10) for shallow-water areas and 0.008 ± 0.007 (n = 10) for mudflats. The paddy fields were not utilized (Fig. 2).
Figure
2.
Habitat utilization rates of wintering Hooded Crane at three wintering stages
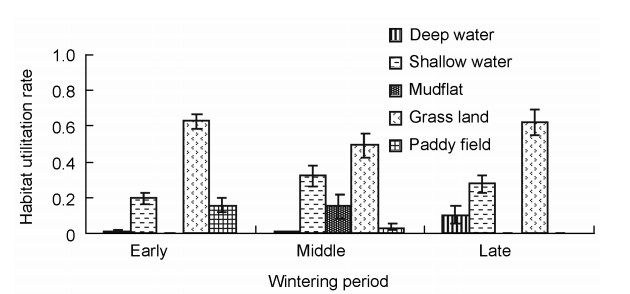
The major habitat types utilized by the goose guild included deep-water and shallow-water areas, mudflats and paddy fields. The grassland habitat was mainly utilized at the early wintering stage, with a utilization rate of 0.627 ± 0.036 (n = 15), followed by shallow-water areas of 0.201 ± 0.033 (n = 15), paddy fields of 0.161 ± 0.038 (n = 15), deep-water areas of 0.009 ± 0.005 (n = 15) and mudflats of 0.001 ± 0.000 (n = 15). At the middle wintering stage, the utilization rate was 0.491 ± 0.069 (n = 13) for grasslands, 0.323 ± 0.059 (n = 13) for shallow-water areas, 0.147 ± 0.069 (n = 13) for mudflats and 0.034 ± 0.018 (n = 13) for paddy fields. The deep-water areas were rarely utilized, with a utilization rate of 0.004 ± 0.004 (n = 13). At the late wintering stage, the major habitat utilized was grassland with a utilization rate of 0.616 ± 0.072 (n = 10), followed by 0.277 ± 0.052 (n = 10) for the shallow-water area and 0.107 ± 0.051 (n = 10) for the deep-water area. The mudflat and paddy field habitats were basically not utilized (Fig. 3).
Figure
3.
Habitat utilization rates of wintering geese at three wintering stages
At the early wintering stage, significant differences for habitat utilization rates between the Hooded Crane and goose guild were found (deep-water: df = 28,
Z = −2.105, p = 0.035; shallow-water: df = 28, t = 3.505,
p = 0.002; mudflat: df = 28, t = −2.931, p = 0.007; paddy field: df = 28, t = −2.686, p = 0.012) in the five types of habitats except for grassland (df = 28, t = 1.849,
p = 0.075). Significant differences were also observed in the utilization rates of shallow-water habitat at the middle wintering stage (df = 24, Z = −2.590, p = 0.010), but not in other habitats (deep-water: df = 24, Z = −1.000,
p = 0.317; mudflat: df = 24, t = −1.690, p = 0.104; grassland: df = 24, t = 0.403, p = 0.690; paddy field: df = 24,
Z = −0.633, p = 0.526). Extremely significant differences were found in the utilization of the deep-water areas, shallow-water areas and grasslands at the late wintering stage (df = 18, Z = −2.796, p = 0.005; df = 18, t = 4.382,
p = 0; df = 18, t = −4.436, p = 0, respectively). No significant difference was found in the utilization of the mudflats (df = 18, Z = −1.000, p = 0.317) (Table 1).
Table
1.
Comparison of habitat utilization rates of Hooded Cranes and goose guild in the winter
Wintering period
Deep water
Shallow water
Mudflat
Grassland
Paddy field
Z
p
Z/t
p
Z/t
p
Z/t
p
Z/t
p
Early n = 30
−2.105
0.035
3.505
0.002
−2.931
0.007
1.849
0.075
−2.686
0.012
Middle n = 26
−1.000
0.317
−2.590
0.010
−1.690
0.104
0.403
0.690
−0.633
0.526
Late n = 20
−2.796
0.005
4.382
0.000
−1.000
0.317
−4.436
0.000
0.000
1.000
Note: Independent sample t test or Mann-Whitney U test, 0.01 ≤ p < 0.05 significant difference, p < 0.01 extremely significant difference.
Width of spatial niche and overlap of Hooded Crane population and goose guild
The width of the spatial niche of Hooded Cranes and goose guild varied during the wintering stage. This niche width of the Hooded Cranes was 1.057 at the early stage, 1.099 at the middle stage and 0.191 at the late stage. The width of the spatial niche was the highest at the middle stage, followed by the early and late stages. The width of the spatial niche of the goose guild was 0.959 at the early stage, 1.133 at the middle stage and 0.893 at the late stage. The width was the highest at the middle stage, followed by the early and late stages (Table 2).
Table
2.
Width of s patial niche and overlap (Oij) of Hooded Cranes and goose guild at three wintering stages
A difference was found in the niche overlap of the Hooded Crane and goose guild at the three wintering stages. The greatest niche overlap was 0.914 found at the late stage, followed by 0.906 at the middle stage and 0.854 at the early stage (Table 2).
Discussion
Characteristics of habitat utilization of Hooded Crane population and goose guild
Waterbirds have to select suitable habitats when facing constantly changing habitats during the winter (Warnock and Takekawa, 1995; Long and Ralph, 2001; Beerens et al., 2011). Resource partitioning is related to its availability in habitats (Kober and Bairlein, 2009). For the waterbirds assembling in flocks, the availability of resources is an important factor affecting flock dynamics (Gawlik, 2002). Therefore, changes in resource availability would cause dynamic changes of the niche of waterbirds (Pearman et al., 2008). When the number of suitable habitats is reduced, the utilization of other habitats will inevitably increase as a compensation for habitat loss (Gerstenberg, 1979; Warnock and Takekawa, 1995; Long and Ralph, 2001). In our study, the variation in water level and seasonal changes of vegetation structure and human activities jointly affected the availability of wetland resources at Shengjin Lake. Thus, the characteristics of habitats and microhabitats would change, further causing dynamic changes in habitat utilization and width of the spatial niche of the Hooded Crane and goose guild.
Habitat and food availability of waterbirds is closely related to water levels (Safran et al., 2000; Zhao et al., 2010). At the middle wintering stage when the Huangpen Sluice was opened for fishing, the water level of Shengjin Lake dropped quickly and large areas of mudflats became exposed. The underground parts of some aquatic plants were readily accessed, which was favorable for both cranes and geese. At the late wintering stage, the water level rose substantially due to the continuous rainfall in the spring, which submerged most of the mudflats and part of the grasslands. The habitats readily utilized by the waterbirds were reduced and the width of the spatial niches was narrowed (Table 2).
The seasonal changes of vegetation structure might induce changes in the strategy of resource utilization on the part of birds (Kushlan, 1981; Thomson and Ferguson, 2007). At the early stage, vegetation flourished in the grasslands. The temperature declined at the middle stage and the withering of plants reduced the availablity of the aboveground parts of plants to waterbirds. Therefore, the habitat utilization of the grasslands by Hooded Cranes and geese was somewhat reduced at the middle stage (Figs. 2, and 3). During the late stage, the vegetation in the grasslands began to germinate, providing favorable foraging habitats for grazing waterbirds.
Grassland reclamation in wetlands, livestock grazing and an enclosure culture resulted in habitat loss, which lowered the resource availability for waterbirds (Wang et al., 2011). Resource-exploiting aquaculture resulted in severe degradation of submerged vegetation, an important food source for waterbirds (Xu et al., 2008). Agricultural activities by farmers usually interfered with the behavior pattern and foraging rate of waterbirds (Luo et al., 2012). Waterbirds foraging in the paddy fields had to face considerable levels of disturbance (Reif et al., 2008). At the early stage, the rice grains were scattered at high density over the newly-harvested fields with few human disturbances. Therefore, the paddy fields were much utilized by the cranes and geese. At the middle stage, human disturbances in the fields, which had been cultivated and changed to wheat fields, increased in intensity. Simultaneously, the middle stage was also a stage for massive fishing. The fishermen usually exploded firecrackers to drive the waterbirds (mainly Phalacrocorax carbo) away from their cage and pond aquaculture, causing the utilization rate to decline in the paddy fields by the cranes and geese. At the late stage, the frequent agricultural activities and dispelling activities may have added to the difficulty of food resource utilization by waterbirds in the paddy field cultivated into winter wheat (Figs. 2 and 3). The density of rice grains scattered in the fields declined as a result of consumption and soil turning (Lee et al., 2001; Amano et al., 2006). If this happened, the paddy field habitats were abandoned gradually by the cranes and geese.
Resource partitioning of Hooded Crane population and goose guild
When the food resources were limited, the waterbirds concentrated within this limited space to search for available food sources, which increased niche overlap and intensified resource partitioning (Kober and Bairlein, 2009). The width of the spatial niche of the Hooded Cranes and goose guild presented similar tendencies of dynamic change in different winter periods (Table 2). This indicates that they had similar requirements for the resources, thereby leading to resource partitioning. The widths of the spatial niche of the goose guild at all stages were all higher than those of the cranes except at the early wintering stage. This shows that the spatial niche of the Hooded Cranes was restricted due to the interspecific competition, compared with geese in a larger group (O'Connor et al., 1975).
Hooded Cranes and geese shared four common habitat types at Shengjin Lake, i.e., shallow-water areas, mudflats, grasslands and paddy fields. Both used the grasslands intensively with subsequent high utilization rates (Figs. 2 and 3), showing that the greatest habitat competition occurred in the grasslands. Significant differences were found in the utilization rates of all habitats at the early and late stages by the cranes and goose guild (Table 1). This suggests the separation in space, especially when the resource was insufficient. It is a result of different patterns of resource utilization by coexisting species (Oksanen, 1987). It has something to do with the feeding behavior of these cranes and geese, which, in turn, is determined by the morphology of birds, such as the length, width and shape of the bill (Kober and Bairlein, 2009; Aplin and Cockburn, 2012). We discovered in our survey that Hooded Cranes foraged by digging out the underground tubers of plants. The geese mainly feed by biting the aboveground parts of plants. There was spatial separation between the cranes and goose guild. Spatial separation reduces the intensity of resource partitioning (González-Solís et al., 2007). At the early winter, besides grassland habitat, the Hooded Cranes also had a high utilization rate in the paddy fields. The grassland with its abundant Carex was the main habitat utilized by the goose species. Only a small number of Bean Geese utilized the paddy fields. Because the Greater White-fronted Goose is a habitat specialist in China, it prefers to graze on short-sward recessional Carex sedge meadows. The Lesser Whitefronted Goose has a short bill and therefore favors grassland where Carex sedges grow (Zhao et al., 2012; Wang et al., 2012). Their number was low and unstable at Shengjin Lake (Cheng et al., 2009; Wang et al. 2012). At the late wintering stage, the water level rose markedly and the temperature increased. As swimming birds, geese prefer a water habitat. But the cranes were mostly concentrated in the grasslands (Figs. 2 and 3, Table 1).
The lowest spatial niche overlap at the early stage might be explained by the fact that the number of waterbirds at the site had not reached its peak. Besides, human disturbances to the paddy fields were few during the early winter, corresponding to high availability. The lake water retreated at the middle wintering stage, exposing a large number of mudflats, providing foraging habitats for the cranes and geese. Significant differences were found in the utilization rates of shallowwater areas but not in other habitats (Table 1). However, the niche overlap was slightly higher than that at the early winter. This is mainly due to the fact that the harvested paddy fields were planted with winter wheat with an increased level of human disturbances, causing a decline in the utilization rate. The maximum spatial niche overlap was found at the late stage (Table 2). This is because most mudflats and part of the grasslands were flooded due to a heavy rainfall during this period. Agricultural activities on the paddy fields, where now the winter wheat grew, had clearly increased and the number of usable habitats was reduced. The cranes and geese could only find their energy supply in a limited number of habitats before further migration. When the resources were abundant, competitive release would strengthen the niche overlap (Martínez, 2004). However, the effect of competition usually occurs after some time (Pearman et al., 2008). Obviously, the high niche overlap of Hooded Cranes and the goose guild in our study cannot be classified into this category. Rather, they competed violently for the limited amount of resources.
Although the cranes and geese have different foraging strategies, they share similar food requirements and wetland resources. The niche overlap was high between them, resulting in severe interspecific competition. The study of the foraging niche overlap is essential between Hooded Crane population and goose guild that will directly imply an important ecological response to the degraded lake.
Acknowledgements
This research was supported by the National Natural Science Foundation of China (Grant No. 31172117), the Graduate Student Innovation Research Projects of Anhui University (Grant No. yqh100118) and the Anhui Academic and Technical Leader Fund. We express our sincere thanks to the staff of Shengjin Lake National Nature Reserve for their assistance in the field work.
Alatalo RV, Gustafsson L, Lindén M, Lundberg A. 1985. Interspecific competition and niche shifts in tits and the goldcrest: an experiment. J Anim Ecol, 54: 977–984.
Amano T, Ushiyama K, Fujita G, Higuchi H. 2006. Foraging patch selection and departure by non-omniscient foragers: a field example in White-fronted geese. Ethology, 112: 544–553.
Anna MC, John W, Jon DM. 2010. Contrasting seasonal survivorship of two migratory songbirds wintering in threatened mangrove forests. Avian Conserv Ecol, 5(1): 2.
Aplin LM, Cockburn A. 2012. Ecological selection and sexual dimorphism in the sooty oystercatcher, Haematopus fuliginosus. Austral Ecol, 37: 248–257.
Beerens JM, Gawlik DE, Herring G, Cook MI. 2011. Dynamic habitat selection by two wading bird species with divergent foraging strategies in a seasonally fluctuating wetland. Auk, 128(4): 651–662.
Both C, Van Turnhout CA, Bijlsma RG, Siepel H, Van Strien AJ, Foppen RP. 2010. Avian population consequences of climate change are most severe for long-distance migrants in seasonal habitats. Proc Roy Soc, 277: 1259–1266.
Burger J, Howe MA, Hahn DC, Chase J. 1977. Effects of tide cycles on habitat selection and habitat partitioning by migrating shorebirds. Auk, 94: 743–758.
Cao L, Zhang Y, Barter M, Lei G. 2010. Anatidae in eastern China during the non-breeding season: geographical distributions and protection status. Biol Conserv, 143: 650–659.
Chen JY, Zhou LZ. 2011. Guild structure of wintering waterbird assemblages in shallow lakes along Yangtze River in Anhui Province, China. Acta Ecol Sin, 31(18): 5323–5331.
Cheng Y, Cao L, Barter M, Xu W, Zhang Y, Zhao M. 2009. Wintering waterbird survey at the Anhui Shengjin Lake National Nature Reserve, China 2008/2009. University of Science and Technology of China Press, Hefei, China.
Davis CA, Smith LM. 2001. Foraging strategies and niche dynamics of coexisting shorebirds at stopover sites in the southern great plains. Auk, 118(2): 484–495.
Fox AD, Cao L, Barter M, Rees EC, Hearn RD, Cong PH, Wang X, Zhang Y, Dou ST, Shao XF. 2008. The functional use of East Dongting Lake, China, by wintering geese. Wildfowl, 58: 3–19.
Gawlik DE. 2002. The effectes of prey availability on the numerical response. Ecol Monogr, 72(3): 329–346.
Gerstenberg RH. 1979. Habitat utilization by wintering and migrating shorebirds on Humboldt Bay, California. Stud Avian Biol, 2: 33–40.
González-Solís J, Croxall JP, Afanasyev V. 2007. Offshore spatial segregation in giant petrels Macronectes spp. : differences between species, sexes and seasons. Aquatic Conserv: Mar Freshw Ecosyst, 17: 22–36.
Howes J, Bakewill D. 1989. Shorebird Studies Manual. AWB Publication, Kuala Lumpur, pp 143–147.
Isacch JP, Darieu CA, Martínez MM. 2005. Food abundance and dietary relationships among migratory shorebirds using grass-lands during the non-breeding season. Waterbirds, 28(2): 238–245.
Ishtiaq F, Javed S, Coulter MC, Rahmani AR. 2010. Resource partitioning in three sympatric species of storks in Keoladeo National Park, India. Waterbirds, 33(1): 41–49.
Jenni DA. 1969. A study of the ecology of four species of herons during the breeding season at Lake Alice, Alachua County, Florida. Ecol Monogr, 39: 245–270.
Jiang HX, Xu WB, Qian FW, Chu GZ. 2007. Impact of habitat evolvement and human disturbance on wintering water birds in Shengjin Lake of Anhui Province, China. Chin J Appl Ecol, 18: 1832–1836.
Kelly JP, Evens JG, Stallcup RW, Wimpfheimer D. 1996. The effects of aquaculture on habitat use by wintering shorebirds. California Fish Game, 82(4): 160–174.
Kober K, Bairlein F. 2009. Habitat choice and niche characteristics under poor food conditions: a study on migratory nearctic shorebirds in the intertidal flats of Brazil. Ardea, 97(1): 31–42.
Krebs CJ. 1989. Ecological Methodology. Harper and Row, New York.
Kushlan JA. 1981. Resource use strategies of wading birds. Wilson Bull, 93(2): 145–163.
Lee WS, Rhim SJ, Park CR. 2001. Habitat use of cranes in Cheolwon Basin, Korea. Korean J Ecol, 24(2): 77–80.
Liu ZY, Xu WB, Wang QS, Shi KC, Xu JS, Yu GQ. 2001. Environmental carrying capacity for over-wintering Hooded Cranes in Shengjin Lake. Resour Environ Yangtze Basin, 10(5): 454–459. (in Chinese)
Long LL, Ralph CJ. 2001. Dynamics of habitat use by shorebirds in estuarine and agricultural habitats in northwestern California. Wilson Bull, 113(1): 41–52.
Luo JM, Wang YJ, Yang F, Liu ZJ. 2012. Effects of human disturbance on the Hooded Crane (Grus monacha) at stopover sites in northeastern China. Chinese Birds, 3: 206–216.
Ma JH, Liu Y, Lei JY. 2006. Methods for Bird Surveys — A Practical Handbook. Bird Watching Society, Hong Kong.
Martínez C. 2004. Food and niche overlap of the Scarlet Ibis and the Yellow-crown Night Heron in a tropical mangrove swamp. Waterbirds, 27(1): 1–8.
Mittelach GG. 1984. Predation and resource partitioning in two sunfishes (Centarchidae sp.). Ecology, 65: 499–513.
Morrison RIG, Davidson NC, Wilson JR. 2007. Survival of the fattest: body stores on migration and survival in red knots Calidris canutus islandica. J Avian Biol, 38: 479–487.
O'Connor RJ, Boaden PJS, Seed R. 1975. Niche breadth in bryozoa as a test of competition theory. Nature, 256: 307–309.
Oksanen L. 1987. Interspecifiic competition and the structure of bird guilds in boreal Europe: the importance of doing fiieldwork in the right season. Trends Ecol Evol, 2: 376–379.
Pearman PB, Guisan A, Broennimann O and Randin CF. 2008. Niche dynamics in space and time. Trends Ecol Evol, 23: 149–158.
Reif J, Voříšek P, Štastný K, Bejček V, Petr J. 2008. Agricultural intensification and farmland birds: new insights from a central European country. Ibis, 150: 596–605.
Reinert HK. 1984. Habitat separation between sympatric snake populations. Ecology, 65: 478–486.
Ribeiro PD, Iribarne OO, Navarro D, Jaureguy L. 2004. Environmental heterogeneity, spatial segregation of prey, and the utilization of southwest Atlantic mudflats by migratory shorebirds. Ibis, 146(4): 672–682.
Safran RJ, Colwell MA, Isola CR, Taft OE. 2000. Foraging site selection by nonbreeding White-faced Ibis. Condor, 102: 211–215.
Thomson RL, Ferguson JWH. 2007. Composition and foraging behaviour of mixed-species flocks in two adjacent African woodland habitats: a spatial and temporal perspective. Ostrich, 78(1): 65–73.
Vahl WK, van der Meer J, Weissing FJ, van Dullemen D, Piersma T. 2005. The mechanisms of interference competition: two experiments on foraging waders. Behav Ecol, 16: 845–855.
Wang QS, Hu XL. 1986. Observation of Hooded Crane wintering in Shengjin Lake. In: Ma YQ (ed) Crane Research and Conservation in China. Heilongjiang Education Press, Harbin, pp 184–189.
Wang X, Fox AD, Cong P, Barter M, Cao L. 2012. Changes in the distribution and abundance of wintering Lesser White-fronted Geese Anser erythropus in eastern China. Bird Conserv International, 22(2): 128.
Wang Y, Fan BL, Ding YR, Pang SS. 2011. The current situation and discussion on wetland ecological restoration of the middle and lower Yangtze River. China Water Res, 13: 4–6.
Warnock SE, Takekawa JY. 1995. Habitat preferences of wintering shorebirds in a temporally changing environment: Western Sandpipers in the San Francisco Bay estuary. Auk, 112(4): 920–931.
Willard DE. 1977. The feeding ecology and behaviour of five species of herons in southeastern New Jersey. Condor, 79: 462–470.
Xiang GE, Wang KF. 2005. Research on the conservation and sustainable utilization of Shengjin Lake wetland resources. Territ Nat Resour Stud, 1: 40–41. (in Chinese)
Xu LL, Xu WB, Sun QY, Zhou ZZ, Shen J, Zhao XX. 2008. Flora and vegetation in Shengjin Lake. J Wuhan Bot Res, 27(3): 264–270.
Yang XL. 2011. Research on the numbers, distribution, feeding behavior and diet of Great White Fronted Geese (Anser albifrons) at Shengjin Lake, the national natural reserves in Anhui Province. Dissertations. University of Science and Technology of China, Hefei, China.
Zhao MJ, Cao L, Fox AD. 2010. Distribution and diet of wintering Tundra Bean Geese Anser fabalis serrirostris at Shengjin Lake, Yangtze River floodplain, China. Wildfowl, 60: 52–63.
Zhao MJ, Cong PH, Barter M, Fox AD, Cao L. 2012. The changing abundance and distribution of Greater White-fronted Geese Anser albifrons in the Yangtze River floodplain: impacts of recent hydrological changes. Bird Conserv Int, 22(2): 135–143.
Zhou B, Zhou LZ, Chen JY, Cheng YQ, Xu WB. 2010. Diurnal time-activity budgets of wintering Hooded Cranes (Grus monacha) in Shengjin Lake, China. Waterbirds, 33(1): 110–115.

 DownLoad:
DownLoad:





 Email Alerts
Email Alerts RSS Feeds
RSS Feeds