Siegfried KLAUS, Winfried VOIGT, Yun FANG, Peter SELSAM, Yue-Hua SUN. 2013: Patch occupancy by the Chinese Grouse (Tetrastes sewerzowi) in a fragmented landscape. Avian Research, 4(3): 232-239. DOI: 10.5122/cbirds.2013.0019
Citation:
Siegfried KLAUS, Winfried VOIGT, Yun FANG, Peter SELSAM, Yue-Hua SUN. 2013: Patch occupancy by the Chinese Grouse (Tetrastes sewerzowi) in a fragmented landscape. Avian Research, 4(3): 232-239. DOI: 10.5122/cbirds.2013.0019
Siegfried KLAUS, Winfried VOIGT, Yun FANG, Peter SELSAM, Yue-Hua SUN. 2013: Patch occupancy by the Chinese Grouse (Tetrastes sewerzowi) in a fragmented landscape. Avian Research, 4(3): 232-239. DOI: 10.5122/cbirds.2013.0019
Citation:
Siegfried KLAUS, Winfried VOIGT, Yun FANG, Peter SELSAM, Yue-Hua SUN. 2013: Patch occupancy by the Chinese Grouse (Tetrastes sewerzowi) in a fragmented landscape. Avian Research, 4(3): 232-239. DOI: 10.5122/cbirds.2013.0019
The Chinese Grouse (Tetrastes sewerzowi) is a rare, endemic bird in China, inhabiting conifer-dominated mountain forests. Both the natural fragmentation and heavy cutting of mature forests have resulted in patchy grouse habitats. We used SPOT (XS-sensor) satellite imagery to discriminate between open land and conifer or broadleaf forests. The area analyzed is about 120000 ha in size and includes the Lianhuashan Nature Reserve and the Yeliguan Forestry Park. We identified 4111 ha of mature coniferous forests in 229 patches (maximum 332 ha, mean 18 ha) as the habitat used by Chinese Grouse throughout the year. We examined 31 forest islands of different sizes and degrees of isolation for the presence of Chinese Grouse. We used generalized linear models (GLM) with binomial error structure and logit link function to estimate the probability of Chinese Grouse occupancy in a forest fragment. Habitat patch size (hs) and distance to the next occupied fragment (doc) were used as predictor variables, important for occupancy. Small habitat islands were disproportionately less likely to be occupied than large, nearby habitats. There was a clear specific habitat size of about 40 ha, above which habitat fragments were occupied more often. Suitable habitat fragments isolated by more than 2 km appeared to be inaccessible to Chinese Grouse. The results have been used in reforestation projects to establish linking corridors in the study area.
Theories of island biogeography (MacArthur and Wilson, 1967) were developed to explain the occurrence of species on islands. The most important variables are island size and isolation from a source area, which also can be applied to habitat fragments in landscape mosaics (Hanski and Gilpin, 1991). The most obvious distinction between real islands and habitat fragments is the matrix, which is water in the first case and other kinds of habitat with varying degrees of unsuitability for a given species in the second case. The matrix may consist of anything except the habitat suitable for the species and is also expected to indicate a more dangerous environment (Stamps et al., 1987). The quality of the matrix is likely to affect the distances an individual moves. More hostile surroundings may reduce the colonization probability (Åberg et al., 1995). An open land matrix could be an isolating barrier blocking any colonization. Many forest bird populations have declined as the result of human-induced habitat fragmentation (Wilcove et al., 1986; Saunders et al., 1991) ― a process of subdividing a continuous habitat into smaller pieces. This process includes the loss of habitat area, reduction of habitat size and an increase in isolation of the habitat patches. One example is the Hazel Grouse (Tetrastesbonasia), which seems to be a poor disperser (Swenson, 1991), whereas to a "good disperser" this kind of landscape may not be regarded as fragmented.
The Chinese Grouse (Tetrastes sewerzowi), a rare and endemic bird in China (category I of the China Red Data Book, "near threatened" in the IUCN Red List), inhabits conifer forests at elevations between 2700 and 3500 m in our study area. The population is thought to be in decline (Sun et al., 2003) because of habitat isolation and fragmentation caused by logging, deforestation, agriculture and other human activities. Remote sensing has been used as a powerful tool for obtaining knowledge about the habitats of endangered species, particularly in large areas that are difficult to access (De Wulf et al., 1988; Homer et al., 1993; Klaus et al., 2001; Jeganathan et al., 2004; Gottschalk et al., 2007). We conducted this study in order to describe the habitats of Chinese Grouse in a larger landscape and to determine the most critical variables (size of habitat patches and distances between occupied patches) as a precondition for habitat linking activities.
Methods
Study area
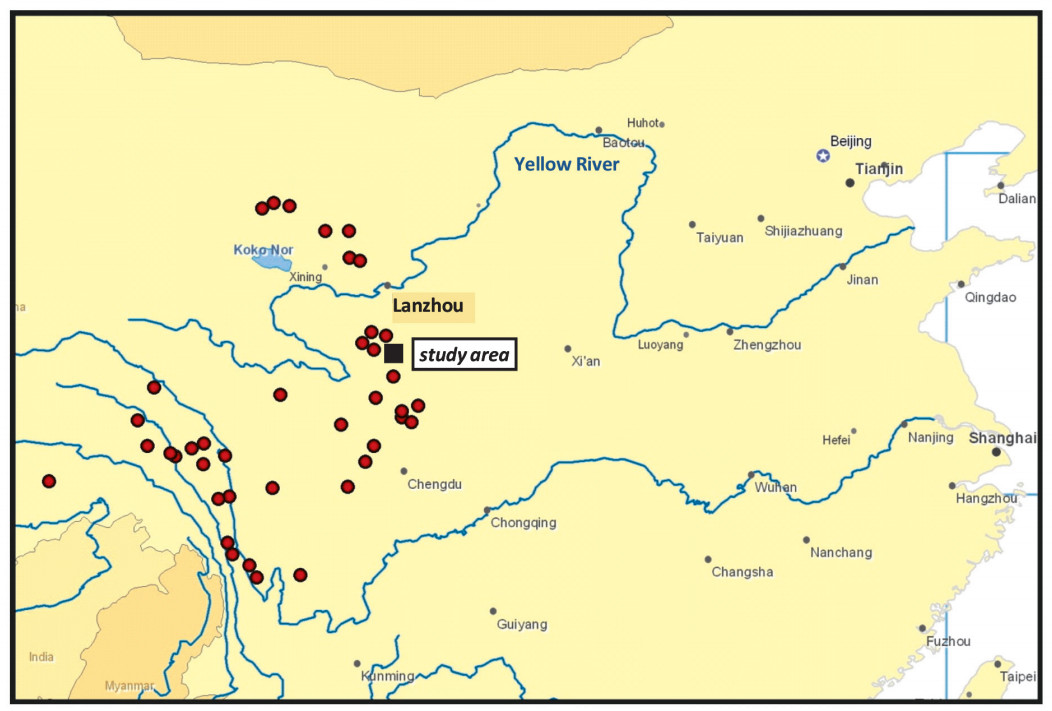
The study area includes the Lianhuashan Nature Reserve (34°40′67″N, 103°30′84″E), a protected area of 11691 ha established in 1983 and the neighboring Yeliguan Forest Park, both in the Lianhuashan mountains, Gansu Province, central China (Fig. 1). The well characterized Lianhuashan Reserve contains selectively cut and uncut mature forests (28%), young forest (10%), shrublands (19%) and grassland and cropland, including villages (43%); its main vegetation types were described by Li and Zhang (1992). Both parts of the study area are separated by a grassland gap of less than 1 km. Shrublands, however, can be extensive. Forests with conifers occur to an elevation from 2700 m up to 3500 m, but grow only rarely on the southerly slopes. The average annual temperature in the reserve is 5.1–6.0 ℃, with extremes of 34 ℃ and −27.1 ℃ in the study area at an elevation of 2100 m. The climate is semiarid and, at the higher elevations, the annual precipitation is about 650 mm. The Lianhuashan Reserve has been described in more detail by Klaus et al. (1996, 2009) and Sun et al. (2003, 2008).
Figure
1.
Distribution (dots) of Chinese Grouse with nominate subspecies north and T. s. secunda south of the Yellow River (Klaus et al., 2009) and location of the study area (square) south of Lanzhou.
The natural fragmentation of forests due to dryness of the southern slopes and heavy cutting of mature forests that occurred prior to reserve establishment have resulted in grouse habitats occurring as patches surrounded by an open matrix (grassland, alpine meadows). The Chinese Grouse population in the Lianhuashan study area is isolated, with the closest known neighboring population 30 km away (Sun et al., 1999, 2006).
Estimation of patch size and distance
We defined a forest fragment as a forest patch surrounded by open land (matrix), usually grassland with or without shrubs, but rarely fields. Earlier studies have shown that the Chinese Grouse habitat is characterized by conifers for cover and willows as the dominant food (Klaus et al., 1996, 2009; Sun et al., 2006). Habitat patches are dominated by conifers, and may contain deciduous forest and/or shrubs as well. Pure deciduous forests, which do not form year-round habitats used by Chinese Grouse, often occur adjacent to a coniferousdominated forest.
We used a SPOT (XS-sensor) satellite imagery (dated 11.09.1998), which allowed the discrimination of open land versus conifer and broadleaf forests. A detailed description of the classification process (software imaging ERDAS on a workstation) has been presented elsewhere (Klaus et al., 2001). The area analyzed was 25% of the SPOT scene, a 30 km × 40 km area (120000 ha). The analysis is based on the precise determination of pixel color (spectral signature).
The classification process resulted in 5 main vegetation types: 1) conifer-dominated forest islands, the preferred habitat for the Chinese Grouse (Klaus et al., 1996), 2) deciduous tree-dominated forest islands, mainly forest succession growing on logged areas (birch Betula albo-sinensis, willow Salix spp., seabuckthorn Hippophae rhamnoides, and other shrubs); without conifers this is not a habitat for Chinese Grouse, 3) moist alpine grassland, 4) dry grassland with scattered shrubs and 5) bare ground (rocks, fields, settlements). The resolution accuracy was tested by terrestrial vegetation analysis combined with photographs showing the distribution of conifer-deciduous trees.
Digitized SPOT satellite images were used to identify the number, size, and location of conifer-containing forest islands, which is the major habitat for Chinese Grouse. The minimum size of a Chinese Grouse territory is 3 ha, based on radio telemetry (Sun and Fang, 1997). Therefore, the results in Table 1 include forest patches > 3 ha up to the largest fragments < 2100 ha. The size and distances between remaining patches were calculated by GIS analysis using ERDAS software. From the resulting working maps (Fig. 2), we selected forest habitat patches that were clearly separated by a distance of > 100 m for checking the occupancy by Chinese Grouse. A survey of Chinese Grouse presence was conducted in May 2000 and 2001, when territorial activity was culminating. The landscape pattern had not changed during the last 20 years, as indicated by recent vegetation studies (Klaus et al., 2009). There was no further logging, and so forest succession on older logged areas proceeds very slowly because of high elevations and grazing by livestock on the open land.
Table
1.
Size distribution of forest fragments dominated by deciduous trees or by conifers
Figure
2.
Example of GIS generated field maps: the dark core area represents conifer-dominated patches. The surrounding ring structures define distances of 100 m each.
Checking forest fragments for the presence of Chinese Grouse
Because resources and accessibility of the study area were extremely limited by the wilderness and remoteness of the mountains (missing roads, steepness of slopes and high elevations > 3500 m a.s.l.), 31 forest patches of different sizes and different distances from the next occupied patch or from the main forest were selected and tested for the presence of Chinese Grouse (Sun et al., 1999, 2006). Because only patches could be selected which were accessible to us and could be visited during the time field work was conducted, we tried to cover a broad range of patch sizes and distances. Grouse were observed during the periods of higher annual and daily activity (morning, evening), when the birds were feeding in willows or displaying. We also recorded indirect signs, such as feathers and excrements. In each fragment we recorded the habitat quality (mixture and/or close vicinity of conifers for cover and willows for food) (Sun et al., 1999). A patch was considered to be occupied where grouse were seen, heard, or indirect signs were found. Indicators of presence and absence of these forest patches were used in our statistical analysis.
Statistics
The research question was: what increases the probability of a habitat being occupied by Chinese Grouse individuals? Habitat patch size (hs) and distance to the next occupied fragment (doc) were used as predictor variables known to be important for the occupation of habitat patches by the grouse. The response variable is binary, for it represents incidences (successful occupations or absence). Because binary responses are strictly bounded to a lower (0) and an upper (1) threshold, the error structure is not normal but binomial. Generalized linear models (GLM) with a binomial error structure and logit link function were used to estimate the probability of Chinese Grouse occupancy in a forest fragment. This class of linear models is preferable to simple linear (multiple) regressions and also to ordinary logistic regression, because it provides more flexibility: the error structure can be specified appropriately as well, since it allows the latitude to select a link function that provides for the lowest deviance. With GLM, the discrepancy between the data and the model is expressed as "deviance". It is equivalent to the sum of squares in simple linear regression. The parameters in GLMs are estimated by maximum likelihood estimation. We started by fitting the full model containing the two explanatory variables: occupancy by grouses ~ hs + doc. Any model simplification by removing stepwise either hs or doc, reduced the explanatory power of the model significantly. Hence, the minimal adequate model is the full model. To compare and simplify models, we used the generic anova function in S-PLUS, which applies a chi-square test. Afterwards, we also tested for nonlinear and interaction effects using an extended model: occupancy by grouses ~ hs + doc + hs2 + doc2 + hs*doc. All terms of the full extended model were significant (p ≤ 0.05), except for hs2 (p = 0.46). Hence, we deleted hs2 as well as doc2 from the model, the latter because there is no sensible interpretation of a quadratic relationship between occupancy by grouse and the size of unoccupied habitat patches. Just as with the previous simpler model, any further simplification by deleting the interaction term resulted in a significant loss in explanatory power, i.e., the current model is appropriate to explain the probability of a habitat being occupied by Chinese Grouse. An initial check for outliers resulted in just one occupied habitat patch with a distance of 5396 m to the next occupied habitat, which is 2.8 times (= 1.5 standard deviation units) more distant than the habitat with the second largest distance (1955 m). We excluded this data point from statistical analyses.
All calculations were done using the package S-PLUS 8.0 (Insightful Corporation, Seattle, USA).
Results
Table 1 summarizes the size distribution of forest fragments of the entire landscape. In the 1200 km2 area, we identified and measured 229 conifer-dominated patches (totaling 4111 ha) and 477 deciduous tree- and shrubdominated forest fragments (totaling 18760 ha). Only eight fragments (3%) were larger than 100 ha. The mean area of all conifer-dominated islands was 18 ± 31.9 ha. Deciduous-dominated forest forms the majority in the landscape. The mean area size of deciduousdominated fragments was 39 ± 199 ha. Due to the large variation in fragment size, the standard deviation was high. This category included the three largest fragments of the study area (1959 ha, 2090 ha and 2103 ha). During the search for Chinese Grouse, we found that this bird can survive at very low density in deciduous-dominated fragments when at least 5%–10% of the conifers were left (Sun et al., 1999).
The mean size of habitat fragments (n = 16) occupied by Chinese Grouse was 102 ± 157 ha (range 5.7–609 ha, median = 46 ha). The mean distance to the next occupied habitat patch was 582 ± 500 m (range 78–1955 m, median = 450 m). The mean size of fragments (n = 15), not occupied by Chinese Grouse was 8.5 ± 11.5 ha (range 3.4–36 ha, median = 3 ha). The smallest forest habitat patch occupied by Chinese Grouse was only 5.7 ha. The matrix was alpine grassland, partially interspersed with tiny (< 40 cm) shrubs of Potentillafruticosa. The minimum significant adequate model we found (p ≤ 0.01) was the full model including an interaction term of the two quantitative explanatory variables in this order:
occupancybygrouse∼hs+doc+hs⋆doc
where hs is the habitat patch size and doc is the distance to the next occupied fragment (see methods). The stepwise model simplifications using the anova function in S-PLUS did not allow us to reduce this model without considerable loss of explanatory power. The larger the habitat islands and the smaller the distance to the next occupied habitat, the higher the probability that a habitat fragment would be occupied by Chinese Grouse. The reduction of deviance by including the three variables in the linear model was considerable; about 80% of the original deviance (from 42.9 to 8.8) in the data (Table 2). hs is more important than doc.
Table
2.
Analysis of deviance table (Binomial model; response: occupancy by grouse) produced by the S-PLUS function anova
df
Deviance
Resid. df
Resid. dev.
Pr(Chi)
NUL
30
42.94286
hs
1
17.33649
29
25.60637
0.00
doc
1
12.78574
28
12.82063
0.00
hs*doc
1
4.05611
27
8.76452
0.04
The terms (predictor variables) have been added sequentially. Pr(Chi): probability obtained by a chi-square test that the reduction in deviance (= goodness of fit) by including a new variable is caused by chance alone (NULL model). The three variables reduce significantly the residual deviance from 42.9 to 8.8.
doc: distance to the next occupied fragment; hs: habitat patch size.
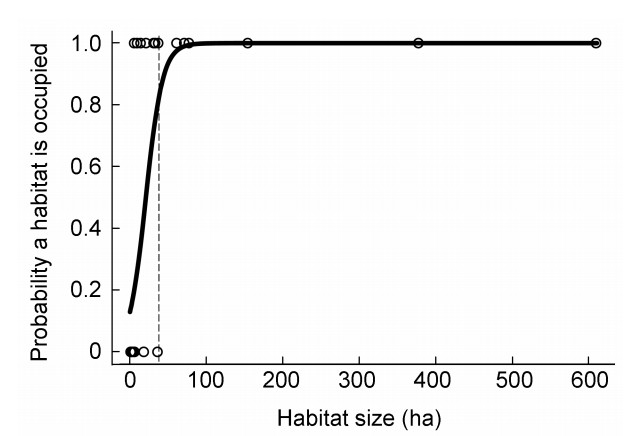
The probability that a habitat fragment was occupied by Chinese Grouse increased with fragment size (Fig. 3). Above a fragment size of 50 ha, the probability a habitat patch being occupied was 1.0. A fragment size of 25 ha corresponded to a probability of occupation of about 0.5 (50%). Habitats larger than denoted by the dotted line (at about 40 ha) were always occupied.
Figure
3.
Relationship (logistic regression response curve) between Chinese Grouse occupancy of fragmented habitats and fragment size (ha) in a fragmented landscape in the Lianhuashan mountains, Gansu Province, China. The dotted line marks the maximum size a habitat may not be occupied. Larger habitats were always occupied.
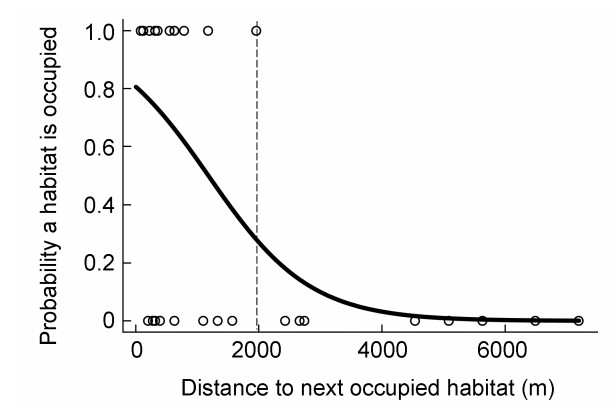
The probability that a habitat fragment was occupied by Chinese Grouse declined with increasing distance to the next occupied fragment (Fig. 4). The 50%-threshold is at about 1000 m. The upper limit appeared to be about 2000 m (dotted line). Habitats outside this range, except for one outlier at 5396 m, were not found to be occupied.
Figure
4.
Relationship (response curve) between Chinese Grouse occupancy of fragmented habitats and their distance (m) to the next occupied habitat in a fragmented landscape in the Lianhuashan mountains, Gansu Province, China. The dotted line marks the maximum distance the grouse are able to cover and eventually occupy a new habitat in a fragmented landscape.
Fragmentation of mountain forests in our study area is a result of old historical land-use practices; dryness of southern slopes, all valleys and lower slopes have been transformed to grassland for grazing animals and intensive clear-cutting during the last decades before the reserve and the Yeliguan Forest Park were established. According to Sun et al. (2003), habitat fragmentation has the potential to affect negatively the Chinese Grouse population in Lianhuashan. The low chick survival and male-biased sex ratio found by Sun et al. (2003) are consistent with models of the effects of habitat fragmentation on capercaillie Tetrao urogallus (Storaas and Wegge, 1987; Kurki et al., 1997; Storaas et al., 1999).
Probability of occupation of forest fragments by Chinese Grouse
Our main interest was to define factors limiting the connectivity of the local population of Chinese Grouse, including the minimum size of habitat fragments that could be occupied and the maximum distances between fragments that would allow the dispersal and colonization of distant forest patches. To occupy a habitat successfully, its size (hs) appeared to be slightly more important than the distance (doc) between habitats. The effect of hs might have emerged even more clearly if we could have included usable information about the shape of these habitat fragments and therefore, about the strength of edge effects into our analysis. Moreover, whether Chinese Grouse occupy a habitat patch, or not, in this landscape clearly depends upon the interaction between hs and doc. This means that there is no simple additive relationship between the two variables and the occupation probability of habitats by Chinese Grouse; the common effect is synergistic (multiplicative). Small isolated habitat islands are disproportionately less likely to be occupied than large, nearby habitats.
Complex, i.e. nonlinear relationships may also occur, because the distances between the habitat islands in our study are measured as linear distances. However, the shortest distance between two suitable habitat patches may not be a straight line, but may depend on various environmental circumstances, such as landscape relief, cover, obstacles, for example small shrubs, lakes and rivers, streets and other urban structures, agricultural areas with hedges, as well as suitable stepping stones.
Based on our data, we found a clear threshold habitat size of about 40 ha, above which fragments were more often occupied (Fig. 3). On the other hand, suitable habitat fragments farther than 2000 m from occupied fragments obviously were inaccessible to Chinese Grouse (Fig. 4). There may be a certain bias, because the habitat patches included in our study were not sampled completely by random selection, so, we cannot fully exclude that there are even larger unoccupied patches.
Our results fit well with the idea of limited colonization of new habitat patches by individual birds of a population depending on the degree of patch isolation (Levins, 1969, 1970), which basically applies the classic theory of island colonization by species (MacArthur and Wilson, 1967) at the population level. It also is in accord with results of similar studies on the closely related Hazel Grouse in Sweden; its occurrence in habitat patches in intensively managed landscapes was also negatively affected by increasing distance between suitable habitat fragments (Swenson 1991; Åberg et al., 1995). The threshold distances for Hazel Grouse were about 200 m in an agricultural landscape and 10 times greater in forests, suggesting a strong effect of the matrix. Hazel Grouse occupancy increased considerably in habitat patches larger than 10 ha (10 ha is a mean territory size for this species in Sweden). Dispersal distances of juvenile Hazel Grouse (September) were 4.8–5.7 km in a forest landscape in NE China (Fang and Sun, 1997). The Chinese Grouse is able to move over larger distances (upper limit 2000 m) in grassland with small shrubs and up to 4 km in forests (Sun and Fang, unpubl.).
Conservation implications
Most of the 229 conifer-dominated patches identified by the remote sensing technique can be regarded as "habitat patches" for the Chinese Grouse, because they are close to each other (open land barrier < 2000 m) and the majority of patches is linked by large tracts of deciduos forest and/or shrubland, allowing dispersal of the grouse in summer. They need special conservation attention. There are two ways to improve the Chinese Grouse habitats on a landscape scale: 1) creating habitat links by conifer plantations in places of large open land barriers and 2) favoring conifer growth by natural succession or planting in deciduous and shrubland patches (both require strict control of livestock grazing in these forests).
Larger habitat patches are most important for the Chinese Grouse and biodiversity in general, because there is room for more territories in a larger patch, stochastic processes are more dramatic to species survival in smaller patches, while predation pressure is higher in landscapes with a higher degree of fragmentation (smaller habitat patches).
In a fragmented landscape, the only way to enable a species to disperse from one patch to the next is to maintain a highly specific connectivity. On the basis of our results, we have suggested to the Forest Administration of Gansu to connect the isolated forest fragments of the Lianhuashan Reserve and the nearby Yeliguan Forest Park and so to increase its connectivity for Chinese Grouse. We also proposed habitat linking by corridor networks with new plantations of conifers between isolated forest fragments within the entire Lianhuashan Mountain range. These suggestions have been accepted and are being implemented. The first plantations have been made, with the objective to form stepping stones and corridors, so that open land barriers will be reduced to < 2000 m.
Acknowledgements
This study was supported by the National Natural Science Foundation of China (Grant No. 31270468, 30620130110), the Chinese Academy of Sciences, the Deutsche Forschungsgemeinschaft (DFG), the BP Conservation programme, the World Pheasant Association. We thank the Administration of Lianhuashan Nature Reserve for continuous support. We are grateful to Hans-Heiner Bergmann, Jon Swenson, Mary F. Gizzie-Voigt and the anonymous reviewer for constructive comments that aided with the revision of the manuscript.
Åberg J, Jansson G, Swenson JE, Angelstam P. 1995. The effect of matrix on the occurrence of hazel grouse (Bonasa bonasia) in isolated habitat fragments. Oecologia, 95: 265–269.
De Wulf RR, Goossens RE, MacKinnon RJ, Cai WS. 1988. Remote sensing for wildlife management: Giant panda habitat mapping from Landsat images. Geocano Int, 1: 41–50.
Fang Y, Sun Y-H. 1997. Brood movement and natal dispersal of hazel grouse Bonasa bonasia at Changbai mountain, Jilin province, China. Wildl Biol, 3: 261–264.
Gottschalk TK, Ekkschmitt K, Isfendiyaroglu S, Gem E, Wolters V. 2007. Assessing the potential distribution of the Caucasian black grouse Tetrao mlokosiewiczi in Turkey through spatial modelling. J Ornithol, 148: 427–437.
Hanski I, Gilpin M. 1991. Metapopulation dynamics: brief history and conceptual domain. Biol J Linn Soc, 42: 3–16.
Homer CG, Edwarts TC, Ramsey RD, Price KP. 1993. Use of remote sensing methods in modelling sage grouse winter habitat. J Wildl Manage, 57: 78–84.
Jeganathan P, Green RE, Norris K, Vogiatzakis IN, Bartsch A, Wotton SR, Bowden CGR, Griffith GH, Pain D, Rahmani AR. 2004. Modelling habitat selection and distribution of the critically endangered Jerdon's couser Rhinoptilus bitorquatus in scrub jungle: an application of a new tracking method. J Appl Ecol, 41: 224–237.
Klaus S, Scherzinger W, Sun Y-H, Swenson JE, Wang J, Fang Y. 2009. Das Chinahaselhuhn Tetrastes sewerzowi Akrobat im Weidengebüsch. Limicola, 23: 1–57.
Klaus S, Scherzinger W, Sun Y-H. 1996. Ökologie und Verhalten des Chinahaselhuhns Bonasa sewerzowi. Der Orn Beobachter, 93: 343–365.
Klaus S, Selsam P, Sun Y-H, Fang Y. 2001. Analyse von Satellitenbildern zum Schutz bedrohter Arten. Fallbeispiel Chinahaselhuhn (Bonasa sewerzowi). Naturschutz und Landschaftsplanung, 33: 281–285.
Kurki S, Helle P, Linden H, Nikula A. 1997. Breeding success of black grouse and capercaillie in relation to mammalian predator densities on two spatial scales. Oikos, 79: 301–310.
Levins R. 1969. Some demographic and genetic consequences of environmental heterogeneity for biological control. Bull Entomol Soc Am, 15: 237–240.
Levins R. 1970. Extinction. In: Gesterhaber M (ed) Some Mathematical Problems in Biology. Am Math Soc, Providence RI, pp 77–107.
Li ZR, Zhang ZH. 1992. Protect and construct Lianhuashan. Jinxing, 6: 8–9 (in Chinese)
MacArthur R, Wilson EO. 1967. The Theory of Island Biogeography. Princeton University Press, Princeton, New Jersey, USA.
Saunders DA, Hobbs RJ, Margules CR. 1991. Biological consequences of ecosystem fragmentation: A review. Conserv Biol, 1: 18–32.
Stamps JA, Buechner M, Krishnan VV. 1987. The effect of edge permeability and habitat geometry on emigration from patches of habitat. Am Nat, 129: 532–552.
Storaas T, Kastdalen L, Wegge P. 1999. Detection of forest grouse by mammalian predators: a possible explanation for high brood losses in fragmented landscapes. Wildl Biol, 5: 187–192.
Storaas T, Wegge P. 1987. Nesting habitats and nest predation in sympatric populations of capercaillie and black grouse. J Wildl Manage, 51: 167–172.
Sun Y-H, Fang Y, Jia C, Wang L, Klaus S, Scherzinger W. 1999. A preliminary study of the effects of habitat isolation and fragmentation on the Chinese grouse at Lianhuashan mountains. Ann Rev WPA, 98/99: 42–48.
Sun Y-H, Fang Y, Klaus S, Martens J, Scherzinger W, Swenson JE. 2008. Nature of the Lianhuashan Naturural Reserve. Liaoning Science Publishing House, Liaoning.
Sun Y-H, Fang Y. 1997. Notes on the natural history and behaviour of the Chinese grouse Bonasa sewerzowi. Wildl Biol, 3: 265–268.
Sun Y-H, Klaus S, Fang Y, Selsam P, Jia CX. 2006. Habitat isolation and fragmentation of the Chinese grouse (Bonasa sewerzowi) at Lianhuashan Mountains, Gansu, China. Acta Zool Sin, 52(Suppl.): 202–204.
Sun Y-H, Swenson JE, Fang Y, Klaus S, Scherzinger W. 2003. Population ecology of the Chinese grouse in a fragmented landscape. Biol Conserv, 110: 177–184.
Swenson JE. 1991. Is the hazel grouse a poor disperser? Trans Int Union Game Biol, 20: 347–352.
Wilcove DS, McLellan CH, Dobson AP. 1986. Habitat fragmentation in the temperate zone. In: Soule ME (ed) Conservation Biology. The Science of Scarcity and Diversity. Sinauer Sunderland, Mass, pp 256–273.
Table
2.
Analysis of deviance table (Binomial model; response: occupancy by grouse) produced by the S-PLUS function anova
df
Deviance
Resid. df
Resid. dev.
Pr(Chi)
NUL
30
42.94286
hs
1
17.33649
29
25.60637
0.00
doc
1
12.78574
28
12.82063
0.00
hs*doc
1
4.05611
27
8.76452
0.04
The terms (predictor variables) have been added sequentially. Pr(Chi): probability obtained by a chi-square test that the reduction in deviance (= goodness of fit) by including a new variable is caused by chance alone (NULL model). The three variables reduce significantly the residual deviance from 42.9 to 8.8.
doc: distance to the next occupied fragment; hs: habitat patch size.

 DownLoad:
DownLoad:






 Email Alerts
Email Alerts RSS Feeds
RSS Feeds