Canchao YANG, Yan CAI, Wei LIANG. 2012: Species identification of sympatric cuckoo nestlings in a multiple-cuckoo system, China. Avian Research, 3(2): 108-112. DOI: 10.5122/cbirds.2012.0016
Citation:
Canchao YANG, Yan CAI, Wei LIANG. 2012: Species identification of sympatric cuckoo nestlings in a multiple-cuckoo system, China. Avian Research, 3(2): 108-112. DOI: 10.5122/cbirds.2012.0016
Canchao YANG, Yan CAI, Wei LIANG. 2012: Species identification of sympatric cuckoo nestlings in a multiple-cuckoo system, China. Avian Research, 3(2): 108-112. DOI: 10.5122/cbirds.2012.0016
Citation:
Canchao YANG, Yan CAI, Wei LIANG. 2012: Species identification of sympatric cuckoo nestlings in a multiple-cuckoo system, China. Avian Research, 3(2): 108-112. DOI: 10.5122/cbirds.2012.0016
Interaction between a parasite and its host could lead to a co-evolutionary arms race. Cuckoo-host system is among the most studied of all brood parasite systems, but the cuckoos of Asia, on the other hand, are much less well known. China has the most abundant cuckoo species in Asia. Many of these co-occur in sympatric areas, posing a potential risk of mis-identification of cuckoo nestlings, especially in Cuculus species. In this study we have provided a practical criterion to identify cuckoo nestlings species in the field and performed molecular phylogeny to confirm our empirical results. These results indicate that two distinct characteristics of cuckoo nestlings, i.e., the gape color pattern and feather traits can be considered as reliable species identification. To our knowledge, this is the first report for species identification of Cuculus nestlings through molecular analysis.
In avian brood parasitism, interaction between a parasite and its host could lead to a co-evolutionary process called arms race, in which evolutionary progress on one side provokes a further response on the other side (Rothstein and Robinson, 1998). The Common Cuckoo (Cuculus canorus) parasitizes more than 100 species of hosts and has evolved more than 20 different kinds of egg phenotypes (Payne, 2005). These usually lay mimetic eggs, which often evolve in improved mimicry under selection pressure of host rejections (Davies, 2000; Yang et al., 2010a). There are up to 17 species of cuckoo species in China (Zheng, 2011; Yang et al., 2012a). Many breed in sympatry, such as the Common Cuckoo, the Himalayan Cuckoo (Cuculus saturatus), the Oriental Cuckoo (C. optatus), the Indian Cuckoo (C. micropterus) and the Lesser Cuckoo (C. poliocephalus) (Yang et al., 2012a). In contrast, only one Cuculus species occurs in Europe. Therefore, there is a potential risk of mis-identification of cuckoo nestlings, especially for the Cuculus species, in a multiple-cuckoo system. Even within a fine-scale study area, several cuckoo species co-occur belonging to the same Cuculus genus. This situation also poses a potential risk in that one host could be utilized by more than one parasite species, or one host parasitized by the same cuckoo species but with polymorphic eggs (Yang et al., 2010a). In this study we demonstrate a practical criterion to identify cuckoo nestlings (Cuculus spp.) in the field. Distinct morphological characteristics of cuckoo nestlings are summarized from our long-term studies and we established a molecular phylogenetic tree of Cuculus species to confirm our empirical results.
Study area
This study was conducted in the Kuankuoshui Nature Reserve, Guizhou Province in south-western China (28°10′N, 107°10′E) during the period from April to July 2004–2011. The study site is situated in a subtropical moist broadleaf and mixed forest, interspersed with abandoned tea plantations, shrub areas and open fields used as cattle pastures. Four Cuculus species, the Common Cuckoo, the Himalayan Cuckoo, the Lesser Cuckoo and the Indian Cuckoo co-occur in our study site (Yang et al., 2010a, 2010b, 2011, 2012a, 2012b). The first three species breed in a sympatric highland area while the Indian Cuckoo breeds in the lowlands.
Methods
Nests of all potential cuckoo hosts were found by systematically searching all typical and potential nest sites and by monitoring the activities of host adults throughout the breeding season. Date of the first eggs laid, egg color, clutch size and occurrence of cuckoo parasitism were recorded and distinct characteristics of different cuckoo nestlings were summarized. Blood samples from cuckoo nestlings were collected late in the brooding period and used for DNA extraction; polymerase cycling assembly (PCA) and sequencing were conducted in the State Key Laboratory of Biocontrol of Sun Yat-sen University. DNA segments from cytochrome b (referred to as cyt b) genes were amplified by using the primer sequences presented in Table 1. Molecular phylogenetic trees were established in MEGA 4.1 by using Neighbor-Joining methods. Another sympatric cuckoo species, the Large Hawk-cuckoo (Hierococcyx sparverioides), whose nestlings are easy to identify (Yang et al., 2012b), was treated as an outgroup for phylogenetic tree construction.
Table
1.
Primer sequences and sources of cytochrome b used in this study
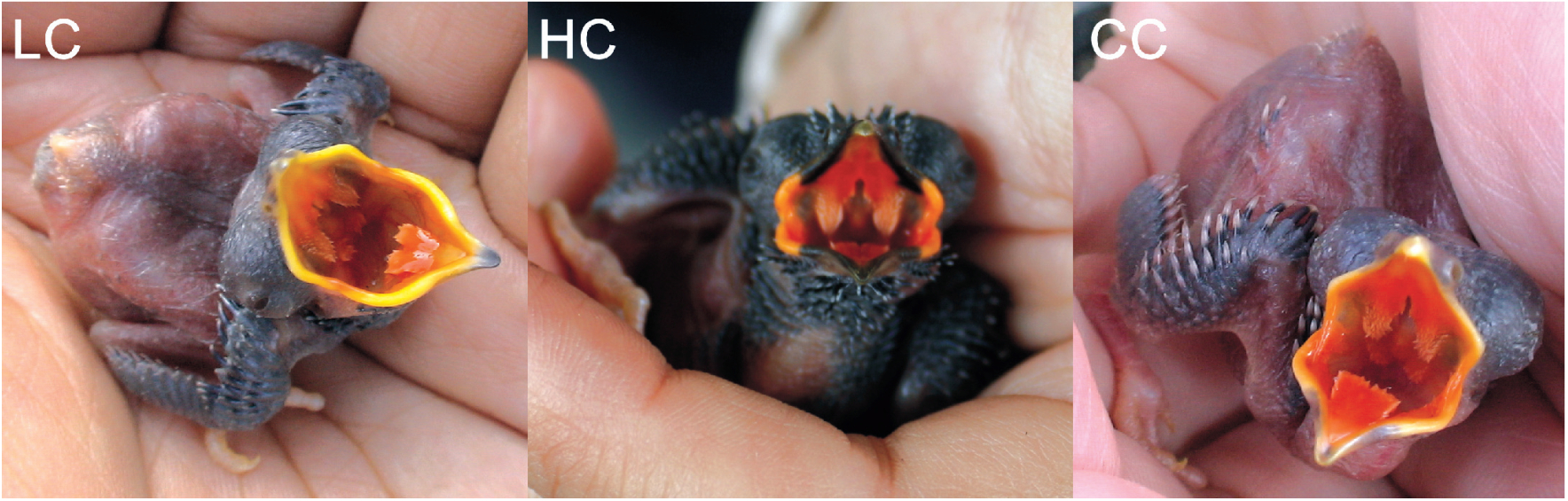
According to our observations and summary of morphological traits, two important and distinct characteristics of cuckoo nestlings can be accepted for reliable species identification. First, the gape color pattern is an apparent characteristic for identification of early stage nestlings, whose skins are naked without plumage. In the Himalayan Cuckoo nestlings (n = 3), two triangular patches on the palate adjoin the cutting edges of the upper mandible; these appeared when the nestlings were four days old and became darker with age and growth (Fig. 1). These black patches on the gapes are absent in other Cuculus nestlings, except for the Oriental Cuckoo, formerly regarded as the same species as the Himalayan Cuckoo (Payne, 2005). Secondly, the plumage color and pattern were treated as important characteristics for identification of cuckoo nestlings at a later stage. Common Cuckoo nestlings (n = 8) have white, narrow tips of feathers on their upper backs and wings, which are obscure in the Himalayan Cuckoo and were absent in nestlings of the Lesser Cuckoo (n = 3) (Fig. 2). The Indian Cuckoo nestlings (n = 4) had wide, large scaled white feather edges, covering their entire head and most of their upper back and wings (Fig. 2). Furthermore, white nape spots only appeared in Common Cuckoo nestlings.
Figure
1.
Photographs of six days old cuckoo nestlings with open gapes. LC refers to the Lesser Cuckoo, HC to the Himalayan Cuckoo and CC to the Common Cuckoo.
Figure
2.
Photographs of near fledged cuckoo nestlings with growing plumage. LC refers to the Lesser Cuckoo, HC to the Himalayan Cuckoo, CC to the Common Cuckoo and IC to the Indian Cuckoo.
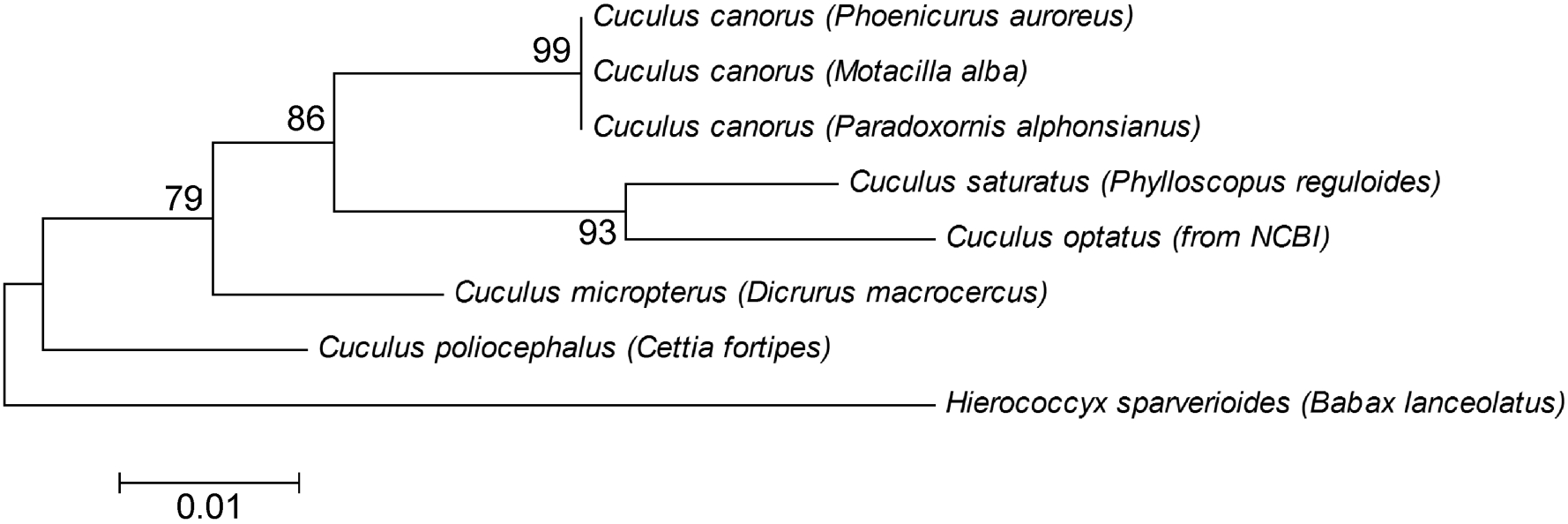
The molecular phylogenetic tree supported our morphological classification of cuckoo nestlings (Fig. 3). Three hosts were found to be used by the Common Cuckoo, i.e., the Daurian Redstart (Phoenicurus auroreus), the White Wagtail (Motacilla alba) and the Ashy-throated Parrotbill (Paradoxomis alphonsianus). As well, the Himalayan Cuckoo, the Indian Cuckoo and the Lesser Cuckoo parasitized Blyth's Leaf Warbler (Phylloscopus reguloides), the Black Drongo (Dicrurus macrocercus) and the Brownish-flanked Bush Warbler (Cettia fortipes), respectively.
Figure
3.
Molecular phylogenetic tree of cytochrome b in Cuculus species by using Neighbor-Joining method. Latin names in brackets refer to the host species, except for the Cuculus optatus, whose sequence was obtained from the NCBI without knowing its host.
In the present study, we demonstrated a practical criterion to identify cuckoo nestlings (Cuculus spp.) in the field. Distinct morphological characteristics of cuckoo nestlings were further confirmed by the molecular phylogenetic tree of these Cuculus species. The cuckoo-host system is among the most studied brood parasite systems, but it should be pointed out that the cuckoos of Asia are much less well known, with the exception of the Japanese cuckoos (Rothstein and Robinson, 1998; Payne, 2005). China has the largest and most abundant occurrence of cuckoo species in Asia, with many of them co-occur in sympatric areas (Yang et al., 2012a). Studies of such multiple-cuckoo systems are needed to reveal host use and competition among sympatric cuckoo species, which can help us to improve our understanding of co-evolutionary processes. However, species identification of cuckoo nestlings is quite difficult in China, especially that of Cuculus species. Even within a fine-scale study area, several cuckoo species co-occur, belonging to the same Cuculus genus. This situation also poses a potential risk that one host could be utilized by more than one parasite species, or one host parasitized by the same cuckoo species but with polymorphic eggs (Yang et al., 2010a). We have provided therefore both a reliable criterion for recognizing morphological traits and a molecular analysis to distinguish different cuckoo nestling species in the field. To our knowledge, this is the first report of species identification of Cuculus nestlings through molecular analysis.
Acknowledgements
We are very grateful to the anonymous referee for constructive comments on an earlier draft of this paper. This work was supported by the National Natural Science Foundation of China (No. 31071938, 31101646), the Key Project of the Chinese Ministry of Education (No. 212136) and by the Program for New Century Excellent Talents in University (NCET-10-0111). We would like to thank the Forestry Department of Guizhou Province and the Kuankuoshui National Nature Reserve for support and permission to carry out this study. We also thank Prof. Zhengwang Zhang for providing photographs and sequences of the Indian Cuckoo, as well as Xinliang Guo, Longwu Wang, Xiao Xu, Na Wang and Tongping Su for assistance with the field work.
Davies NB. 2000. Cuckoos, Cowbirds and Other Cheats. Poyser, London.
Payne RB. 2005. The Cuckoos. Oxford University Press, New York.
Rothstein SI, Robinson SK. 1998. Parasitic Birds and Their Hosts. Oxford University Press, New York.
Yang C, Antonov A, Cai Y, Stokke BG, Moksnes A, Røskaft E, Liang W. 2012b. Large Hawk-cuckoo Hierococcyx sparverioides parasitism on the Chinese Babax Babax lanceolatus may be an evolutionarily recent host-parasite system. Ibis, 154: 200–204.
Yang C, Cai Y, Liang W. 2011. Visual modeling reveals cryptic aspect in egg mimicry of Himalayan Cuckoo (Cuculus saturatus) on its host Blyth's Leaf Warbler (Phylloscopus reguloides). Zool Res, 32: 451–455.
Yang C, Cai Y, Liang W. 2010b. Brood parasitism and egg mimicry on Brownish-flanked Bush Warbler (Cettia fortipes) by Lesser Cuckoo (Cuculus poliocephalus). Zool Res, 31: 555–560. (in Chinese)
Yang C, Liang W, Antonov A, Cai Y, Stokke BG, Fossøy F, Moksnes A, Røskaft E. 2012a. The diversity of parasitic cuckoos and their hosts in China. Chinese Birds, 3: 9–32.
Yang C, Liang W, Cai Y, Shi S, Takasu F, Møller AP, Antonov A, Fossøy F, Moksnes A, Røskaft E, Stokke BG. 2010a. Coevolution in action: disruptive selection on egg colour in an avian brood parasite and its host. PLoS ONE, 5(5): e10816.
Zheng G. 2011. A Checklist on the Classification and Distribution of the Birds of China. 2nd edn. Science Press, Beijing.

 DownLoad:
DownLoad:






 Email Alerts
Email Alerts RSS Feeds
RSS Feeds