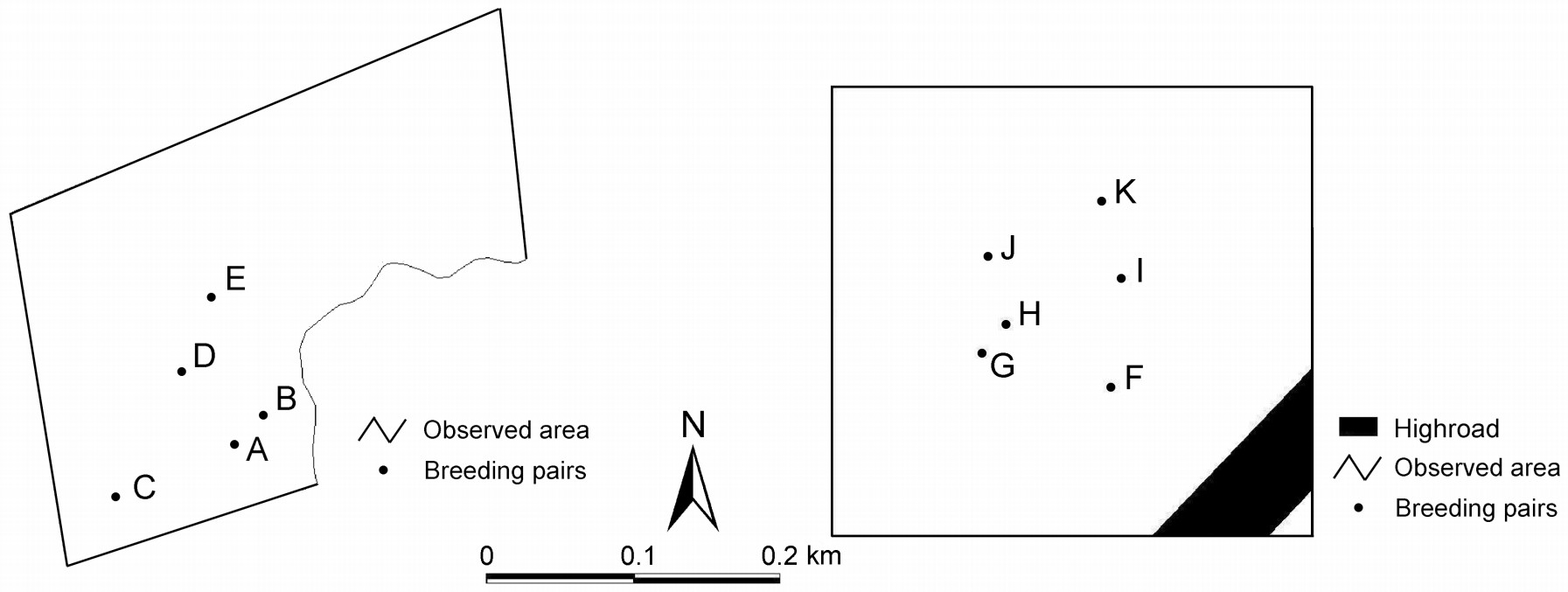
Figure
1.
Observed breeding pairs in Anbanghe and Daqing Nature Reserves
| Citation: |
Weiwei ZHANG, Jianzhang MA. 2012: Use of display platforms of Common Coots (Fulica atra) compared with the use of nests during breeding seasons. Avian Research, 3(2): 82-90. DOI: 10.5122/cbirds.2012.0013

|
We investigated a breeding population of Common Coots (Fulica atra) from April to June in 2008 and 2009. We found that each pair of coots built display platforms beside their nest in their territory during the breeding season. On average, 3.1 (range 2-7) platforms were built by each pair of coots. Display platforms were simpler with less coverage and were significantly lower above the water surface than nests; however, their depth below the water surface was not significantly different. Platforms were primarily used for roosting, preening and mating in the breeding season, but they were also used to store animal food. In the breeding season, nest was usually built later than platforms, the use ratio (total and daily frequency) of platforms decreased after nest was built, and more mating behavior was performed on the nest than on the platform. We compared the use ratio of both sexes to the nest and platforms in different breeding phases; the results show that females used the nest more than the platforms, while males used the platforms more than the females, suggesting that females pay more attention to breeding, while males put more energy in territorial defense. Since coots are territorial during the breeding season, the platforms may play an important role in confirming and recognizing territorial boundaries.
Display platform construction is a well-known behavior of nesting coots (Fredrickson, 1970). The platforms of coots are always built before the nests in the spring, made up from drift wood, sticks or broken-down cattails, particularly the stems and leaves (Gullion, 1954). Platforms are not used for egg laying but instead for chick brooding and roosting (Rek, 2010). Gullion (1954) described American Coots (Fulica americana) using these structures for roosting and preening.
The Common Coot (Fulica atra) is an abundant aquatic bird that is highly territorial during the breeding season, especially in non-migrating populations (Gullion, 1953; Cavé and Perdeck, 1989; Rizi et al., 1999; Chen et al., 2010; Zhang et al., 2011). However, territory formation and defense can be difficult because of the number and proximity of nests constructed in relatively small areas. Previous studies have focused primarily on changes in territorial behavior during the year and the size of territory (Gullion, 1953), the effect of age of individual birds and that of their neighbors on size of territory and its quality (Cavé and Perdeck, , 1989), the effect of age and sex on defense of territory (Ryan and Dinsmore, 1979, 1980) and breeding and guarding behavior (Samraoui and Samraoui, 2007; Cheng et al., 2010; Zhang et al., 2011; Zhang and Ma, 2011).
Building and maintenance of platforms both incur costs in the form of energy for coots during the breeding season. In the present study, we aimed to answer the following questions. Why do coots spend so much time and energy on platforms during the breeding season? How do coots allocate their time during the day between platforms and nest? What role do platforms play in breeding?
We monitored two breeding populations of Common Coots during the early breeding period, at the Anbanghe Nature Reserve (46°53′–47°03′N, 131°06′–131°32′E) and the Daqing Longfeng Nature Reserve (46°28′–46°32′N, 125°07′–125°15′E) in Heilongjiang Province, China. Both are shallow water wetlands where the dominant plants include reeds (Phragmites australis), cattails (Typha angustifolia), broadleaf cattails (Typha latifolia) and calamus (Acorus calamus). The annual temperature of the study area is 2.5–2.6℃ in the Anbanghe wetland and 4.5℃ in the Longfeng wetland. There are thousands of coots in both the Anbanghe and Longfeng wetlands during the breeding season, where coots face human disturbance caused by ecotourism. The population in the Longfeng wetland is also disturbed by speeding vehicles on the road which passes through the entire wetland.
We observed 11 pairs of coots using binoculars in May of 2008 and 2009 (Fig. 1). We used all-occurrence and focal animal sampling methods. The sex of individuals was identified by their call following the description of the American Coot (Gullion, 1950, 1951; Fredrickson, 1968; Eddleman and Knopf, 1985), as well as by their body size, territorial, sex and other behavior. We noticed that the frontal shield of males is larger and wider than that of females and the convex arc of the frontal shield of the male is more forward than that of the female. Hence, we used the frontal shield characteristics to distinguish between the sexes.
We selected pairs whose territories were near lake shores or ponds because their movements are easier to observe. These breeding pairs were less than 200 m away from our observation sites; there was no evidence that as observers we disturbed normal behavior. Observations were made in May when the vegetation (i.e., reeds and cattails) was short and sparse. We also checked the platforms and nests when we searched for the nest structures of coots and surveyed them if there were any traces of coots, such as feathers and faeces. The height above the water surface of the nests and platforms and their depth below the water surface were measured. We classified the vegetation coverage of nests and platforms in 1 m × 1 m plots into eleven categories from 0 to > 90%. Daily observations were made from 05:00–17:00 hours. The breeding phase of coots was referred to as "nest building" (breeding pairs build their nest or platforms, while no eggs are produced), "egg laying" (breeding pairs begin to lay eggs but do not finish the entire procedure) and "incubation" (breeding pairs finish egg laying and begin to incubate them). Observations ended before egg hatching started in June when it became difficult to follow the movements of coots. Thus, our study mainly focused on the early phases of the breeding season. The lengths of the observation time for each pair of coots are shown in Table 1. The total observation time per day for each pair was not always the same. We used the percentage of time spend for comparative purposes.
| Study site | Code of breeding pair | Length of observation (h) | Number of platforms | Breeding period |
| Anbanghe | A | 94 | 3 | Nest building |
| Anbanghe | B | 94 | 2 | Nest building |
| Anbanghe | C | 93 | 3 | Nest building |
| Anbanghe | D | 92 | 5 | Nest building |
| Anbanghe | E* | 90 | 4 | Egg laying |
| Longfeng | F | 93 | 4 | Egg laying |
| Longfeng | G | 60 | 7 | Incubation |
| Longfeng | H | 58 | 3 | Incubation |
| Longfeng | I | 58 | 2 | Incubation |
| Longfeng | J* | 40 | ≥2 | Incubation |
| Longfeng | K* | 40 | ≥2 | Incubation |
| * We could not determine all activities of E, J and K because only part of their platforms could be observed. | ||||
 DownLoad:
CSV
DownLoad:
CSV
All structures built by coots in which eggs were not laid were regarded as display platforms while structures chosen for egg laying were considered nests. Given that the time used for observation of each of the 11 pairs was not the same, we used the relative amounts of time spent on nests and platforms in each day in our statistical analyses.
The statistical package SPSS 17.0 was used for testing whether the data were normally distributed, using the Kolmogorov-Smirnov test and t-tests for comparing differences. Figures were made using SigmaPlot 10.0. Data are presented as mean ± standard error (SE).
The pairs chosen for observation and their breeding phases are shown in Table 1. During the breeding season, the coots built display platforms in their territories before and after nest building.
Each pair of coots built at least two platforms in their territory (Table 1) but usually they built three or four. The largest number of platforms built by the same pair was seven. In a small pond of Anbanghe, where most nests and platforms were likely found, there were 17 nests and 53 platforms (average 3.1 platforms per nest). These platforms were not built at the same time. Usually platforms were constructed before nests, but these structures were also added to territories during the incubation period.
Typically one or two ramps (sometimes three or four) were built on the side of nests, but not on the sides of platforms. Cup-like structures found in the upper part of nests were absent from platforms.
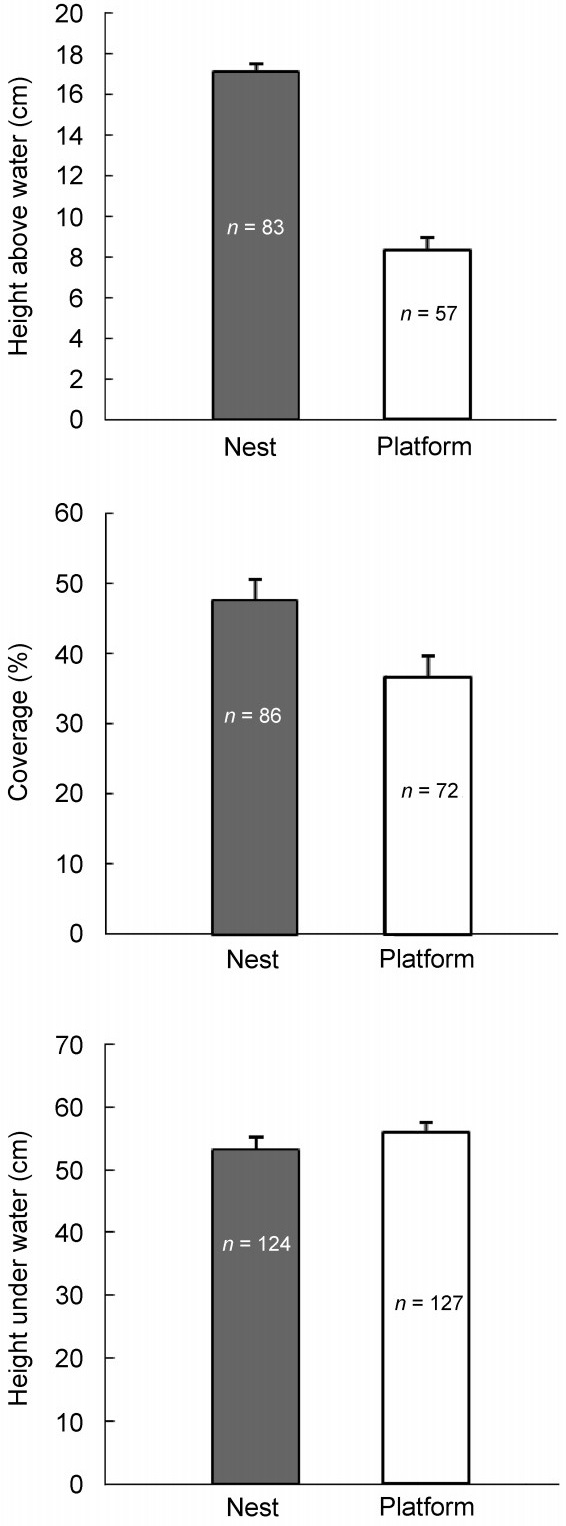
Because the shape of platforms was irregular, compared with that of nests, we compared three basic parameters: heights above and under the water surface and vegetation coverage in 1 m × 1 m plots (Fig. 2). The mean height of nests was 17.1 ± 0.4 cm (n = 83) and that of platforms 8.4 ± 0.6 cm (n = 57), which were significantly different (t = 11.802, df = 138, p < 0.001). However, there was considerable variation in the height of platforms; some were water-soaked, whereas others were as high as the nests.
Figure 2 also shows vegetation coverage and height and depth of nests and platforms with respect to the water surface. The distances of nests and platforms below the water surface did not differ significantly (t = 1.057, df = 249, p = 0.291), but nests had greater vegetation coverage (t = –2.617, df = 156, p = 0.01) (Fig. 2).
Only breeding pairs built platforms in their territory, which they began to build after pair formation. Our observations suggest that platforms are mainly used for roosting and preening, copulation, feeding and as defecation sites, as well as markers of territorial boundaries.
Coots stood or sat on platforms after feeding, washing and sometimes after fighting with neighbors. The number of platforms containing feathers was twice that of nests (39 platforms vs. 14 nests). Of the 14 nests with feathers, only four still had eggs that were not hatched. The platforms or nests were not the only locations for roosting and preening; we also observed these activities in bulrushes or cattails and on driftwood.
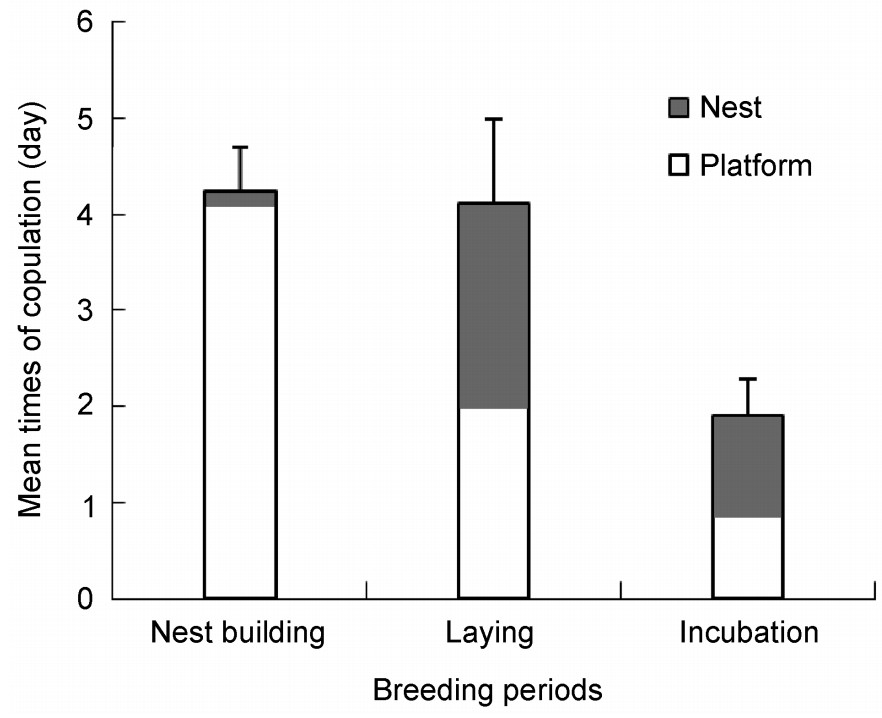
During the breeding season, coots copulated soon after pair formation. Before the nests were built, almost all copulation rituals occurred on the platforms. Following nest construction, the females began to lay eggs and both the nest and platforms were used for copulation during this period. Consequently, the number of copulations on the platforms was fewer than before, but still higher than that during the incubation period (Fig. 3). Differences in the number of copulations/day among the different phases of the breeding period were statistically significant (F2, 49 = 4.564, p = 0.015). Copulation was more frequent before, rather than after the laying of the full clutch (Fig. 3). During the nest building period there were 4.3 ± 0.4 copulations/day (n = 32). Copulations were as frequent before the nests were built as during the nest building period (4.1 ± 0.9, n = 10), when females began to lay eggs. During this period, about half of these activities occurred on the nests. When the females began to incubate the eggs, the number of copulations dramatically decreased to 1.9 per day (SE = 0.376, n = 15); but platforms continued to be used as frequently as nests for mating.
Faeces were found on 38 structures, consisting of seven nests in which the eggs had all hatched, one nest with eggs (the coot was frightened by our appearance and excreted before escaping) and on 30 platforms. Thus, the proportion of platforms with faeces (78.9%) was much higher than the proportion of nests without faeces (21.1%).
We also found elytra of predaceous diving beetles, small fish, residues of frogs and other animal parts on the platforms and nests after the broods had hatched.
Activities on platforms were also associated with territoriality. Some pairs (A, C, D, F and G) were observed adding new platforms when their activities moved from one part to another part of their territory or after fighting with neighbors. Platforms were always built near territorial boundaries. After conflicts with neighbors, one or both sides built platforms near the point of conflict, suggesting they function as a marker of territorial boundaries.
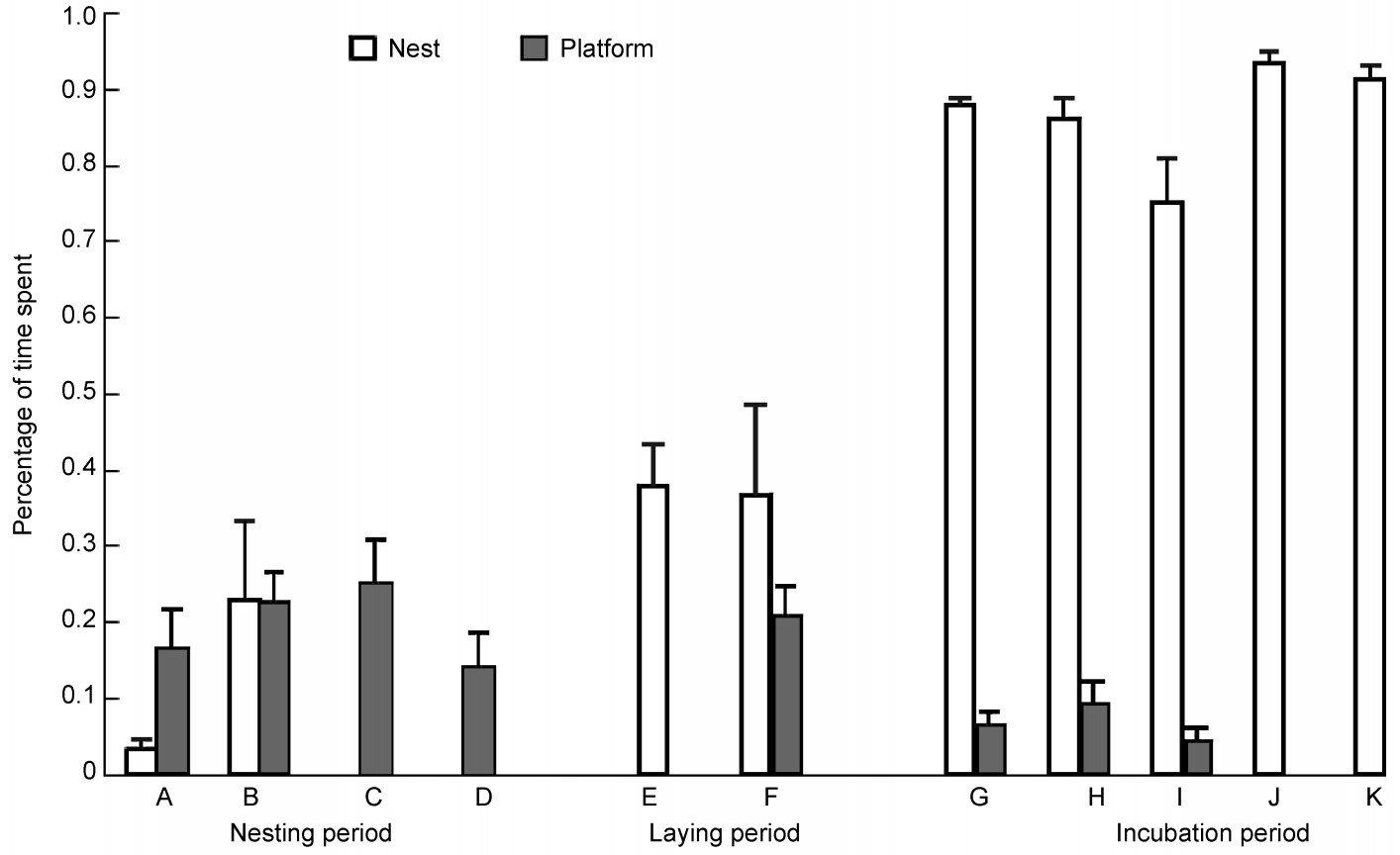
During the nest building period, the percentage of time spent on nests and platforms during the day was 10.8 ± 4.7% (n = 13) and 19.8 ± 2.5% (n = 31), respectively. Statistically, this difference was not significant (t = –1.931, df = 42, p = 0.073). During the egg laying period, the percentage of time spent on their nest each day was 37.3 ± 5.5% (n = 13) and that on platforms 20.7 ± 4.0 (n = 5), which, again, was not significantly different either (t = 1.786, df = 16, p = 0.093). However, during the incubation period, this percentage increased to 86.9 ± 1.8% (n = 25) for nests and decreased to 6.6 ± 1.3 (n = 15) for the platforms; these differences were statistically significant (t = 31.197, df = 38, p < 0.001).
Time spent on the nests dramatically increased from the nest building period to the incubation period (Fig. 4), which was associated with a sharp decline of time spent on the platforms (Fig. 4). Before laying eggs, less than 20% of the time was spent on their future nest; the percentage of time present on the platforms was similar or higher than that on the nest. After the coots began to lay eggs, time in the nests increased to 40%, while time spent on platforms decreased. During the incubation period, time spent on nests was greater than 80% but time on the platforms decreased to less than 10% (Fig. 4).
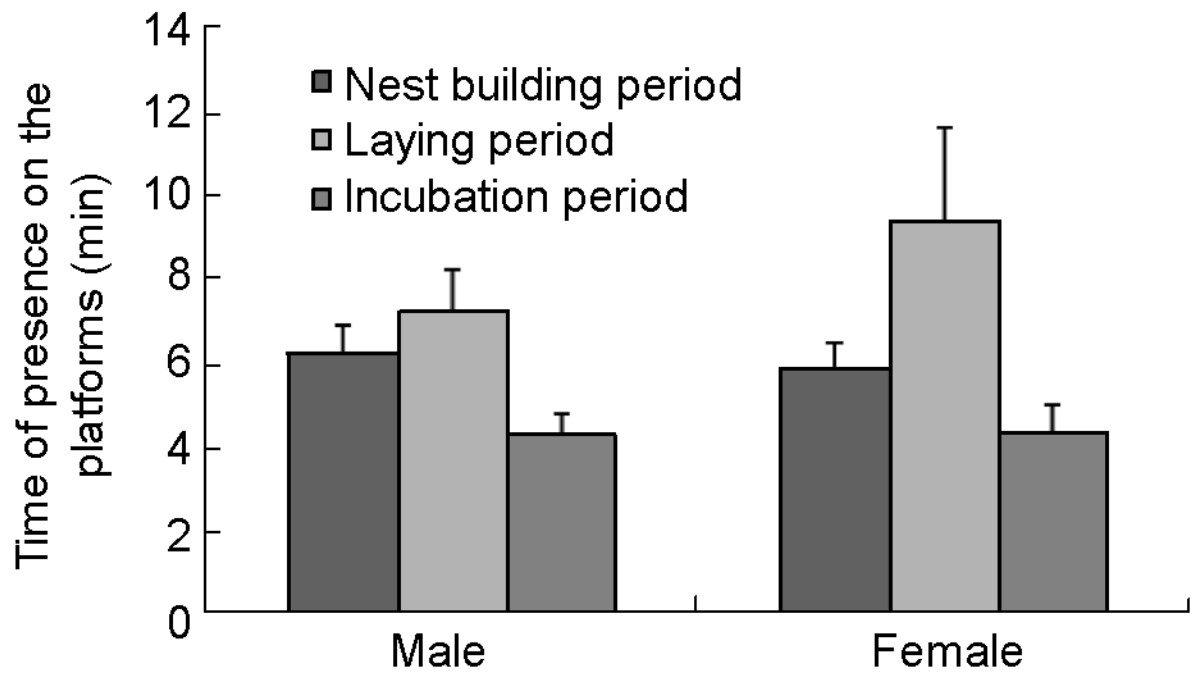
In the nest building period, both sexes frequently used their platforms, where the time spent was similar (Fig. 5, male = 6.3 ± 0.6 min, n = 93; female = 5.9 ± 0. 6 min, n = 63). The difference between the sexes was, statistically, not significant (U = 2830.00, Z = –0.362, p = 0.717). In the egg laying period, the period of presence on the platforms by males was 7.2 ± 1.0 min (n = 29) and that of females 9.4 ± 2.2 min (n = 19). There was no significant difference between the sexes (t = 0.989, df = 46, p = 0.328). When the birds began to incubate, both males and females decreased their period of presence on the platform to 4.3 ± 0.5 min (n = 72) and 4.3 ± 0.6 min (n = 28), respectively. There was also no significant difference between the sexes (U = 841.50, Z = –0.127, p = 0.889).
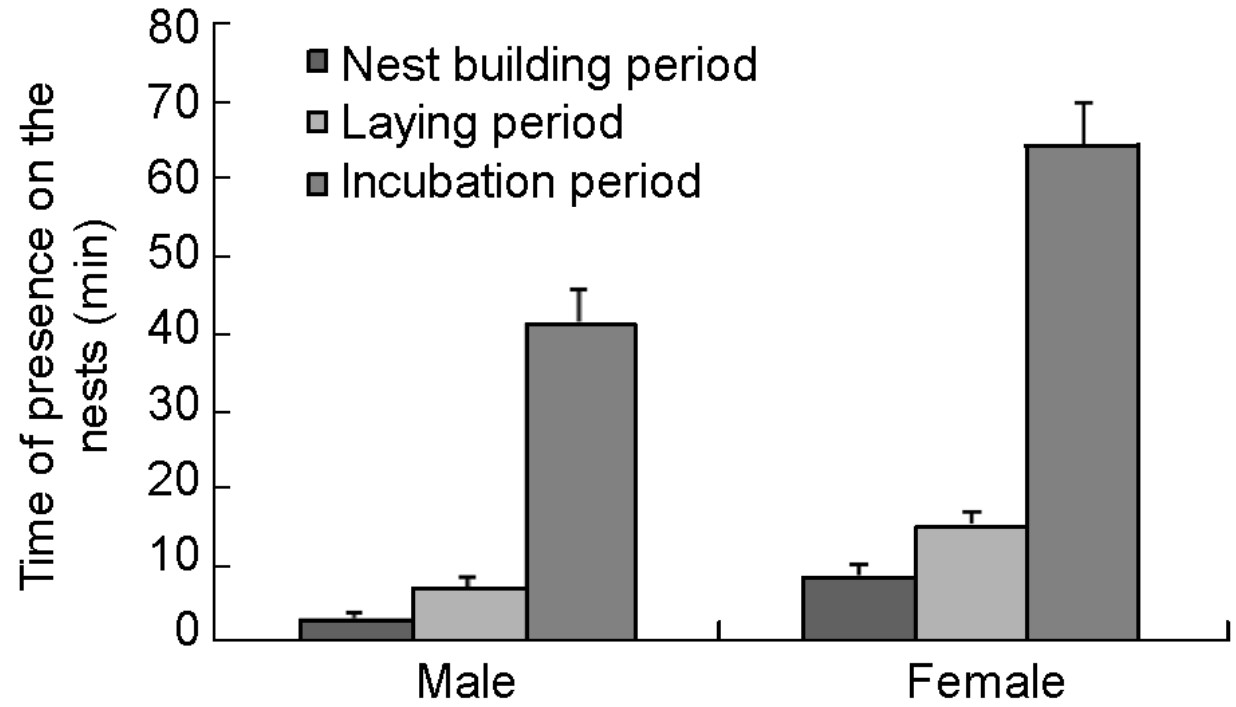
However, in the nest building period, the length of each time females spent on their nests was higher than that of males (males: 3.4 ± 0.6 min/time (n = 72); females: 8.8 ± 1.6 min (n = 28), Fig. 6); the difference was significant (U = 453.00, Z = –4.490, p < 0.001). During the egg laying period, females stayed longer on their nests each time, compared with males (females: 15.5 ± 1.6 min/time, n = 44; males: 7.5 ± 1.1 min, n = 49); this difference was also significant (U = 530.00, Z = –4.429, p < 0.001). Both sexes incubate eggs; however, females took more time in sitting than males in each day (Fig. 6). Females incubated in the nest for 30–136 min each time (mean: 64.1 ± 65.5, n = 21), significantly longer than males, whose time ranged from 3 to 74 min (mean: 41.4 ± 4.2 min, n = 22; t = 3.316, df = 41, p = 0.002).
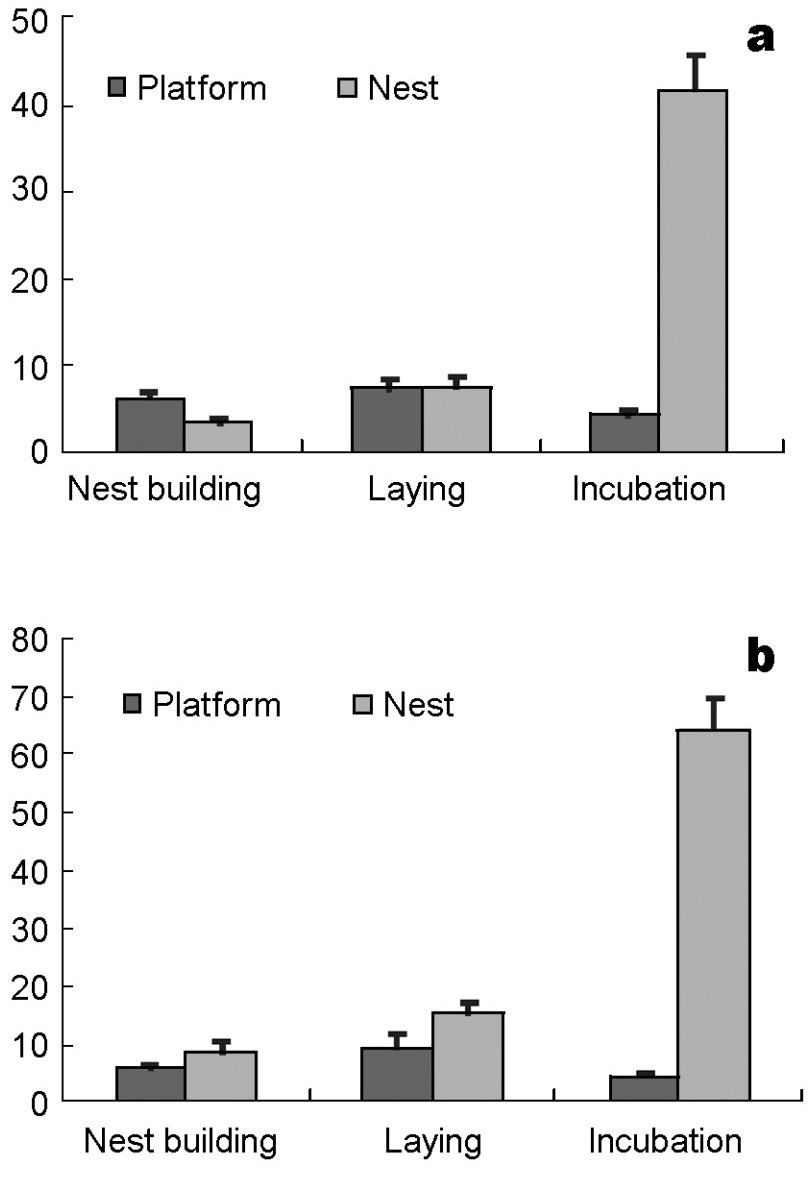
In the nest building period, each time a male was on the platform, the length of his presence was higher than on that the nest (U = 1997.50, Z = –2.726, p = 0.006, Fig. 7a). This was different from females, who showed no significant preference for either the nest or the platforms during this period (U = 735.00, Z = –1.270, p = 0.204, Fig. 7b). During egg-laying, each time the female was on the nest the period of her presence was significantly longer than that on the platforms (t = –2.093, df = 61, p = 0.041); however, the males showed no obvious preference to nest or platforms (U = 640.50, Z = –0.727, p = 0.467). Both sexes stayed much longer on the nest than on the platforms (male: U = 33.50, Z = –6.604, p < 0.001; female: t = 10.813, df = 20.539, p < 0.001) during incubation.
Animal behavior can be thought of as the outcome of a series of trade-offs between costs and benefits associated with performing various actions (Morrell, 2004). This trade-off theory has been broadly used to deepen our understanding of animal behavior (Hutchings et al., 2006). The behavior of coots in platform building during the breeding season is in common with that of other birds. Platforms are indispensable structures. Our observations suggest that the characteristics and role of platforms are different from those of the nest.
Both nests and platforms were built on the water surface, mainly using reeds and cattails. However, platforms were simpler and had less vegetation than nests. Nests were larger than platforms. Platforms were generally similar to nests except for the absence of eggs. Nests containing eggs, were additional structures or built on the foundations of one of the platforms. When the platforms were not used for egg laying, the site selection was not as covert as for nests and much more simpler than nests. Platforms were also used by the coots after the nests were built, but time spent on them dramatically decreased.
Rek (2010) suggested that adult coots build nest-like platforms for chick brooding and roosting. However, we focused on the function of platforms before the chicks hatched. Coots use platforms like a nest without eggs during the breeding season for roosting, preening and copulation, especially before the nests are built. We also suggest that platforms are used to store food that could not be eaten immediately or to eat food that was difficult to catch to avoid the prey from escaping. When the nestlings could not sufficiently feed themselves, the parents would sometimes feed them on the platforms and nests.
Both females and males used their platforms while they took part in defending their territories and patrolling during their entire breeding periods and both used and defended the platforms throughout these periods. However, in various periods, both sexes were present on the two kinds of structures but at different times and frequencies. Compared with females, males spent less time on their nests, especially when they are hatching. Before the females laid eggs, both sexes paid attention largely to building their nests and defending their territory. The differences between the sexes in the presence times at their nest and the total time spent each day there were statistically not significant. They patrolled the nest and platforms, displaying and calling. The time both males and females spent on platforms were similar.
The males appeared more frequently and spent longer time every day on the platforms than the females during breeding periods. In total, males spent less time on the nests than females, suggesting that males put more efforts in building and guarding of the nest than females. Males played more important roles in establishing and defending their territories, while contracting females put more effort into egge production.
The functions mentioned earlier do not explain why coots build several platforms in their territory. In fact, platforms seemed to be substitute structures for the coots. Cut reeds, laid cattails and other floating plants on the water surface could be alternative sites, which can be obtained easily without putting much energy into them (Fredrickson, 1970). Even if platforms were necessary for their behavior, one for each pair should be sufficient. However, the number of platforms was quite large; some breeding pairs built seven platforms, from which we infer that platforms must have another role for coots during breeding.
Wright and Taylor (2000) suggested that nesting platforms might increase aggressive interactions in Common Loons (Gavia immer). Mager III et al. (2008) also found that nest platforms increased aggressive behavior in Common Loons. These results suggest that platforms are associated with territorial behavior. Display behavior can be used by animals experiencing intense competition (Petrie, 1988). The front shield of a coot is used for visual display (Gullion, 1952). We believe that platforms are another form of display, indicating ownership of the surrounding area, along with calls made when entering and leaving the territory. In addition, roosting and preening behavior on the platforms also display ownership.
Nest building is an important part of the reproductive cycle of birds. Nests are built for breeding, unlike platforms which are not directly used for this purpose. During the breeding season, coots defending their territories built platforms, usually following conflict with neighbors. Thus, platform construction seems to be associated with defense of territory. We suggest that platform building is a consequence of territoriality. Gullion (1953) considered that breaks in a cattail edge, willow trees along a shore-line and a stretch of shallow water constituted boundary reference points in a population of American Coots. However, such landscape characteristics seem to be only available for pairs on the edge of a population, hence we conclude that platforms serve as landmarks of territorial boundaries.
This research was supported by the National Natural Science Foundation of China (No. 200904012) and the Jiangxi Agricultural University Research Start Fund for Doctor (09003274). We thank the administration of the Anbanghe Nature Reserve for permission to conduct this research. We are grateful to Yue Zhang and Zhipeng Xu for providing us with their kind help during the field work.
|
Cavé AJ, Perdeck AC. 1989. Size and quality of the coot Fulica atra territory in relation to age of its tenants and neighbors. Ardea, 77: 87–98.
|
|
Cheng K, Ma JZ, Li JB, Jiang R. 2010. Nest site and territory characteristics of the Common Coot (Fulica atra) in Anbang River Nature Reserve, Heilongjiang. Sichuan J Zool, 29(3): 372–376. (in Chinese)
|
|
Eddleman WR, Knopf FL. 1985. Determining age and sex of American Coots. J Field Ornithol, 56(1): 41–55.
|
|
Fredrickson LH. 1970. Breeding biology of American Coots in Iowa. Wilson Bull, 82(4): 445–457.
|
|
Gullion GW. 1951. The frontal shield of the American Coot. Wilson Bull, 63: 157–166.
|
|
Gullion GW. 1952. The displays and calls of the American coot. Wilson J Ornithol, 64(2): 83–97.
|
|
Rizi H, Benyacoub S, Chabi Y, Bańbura J. 1999. Nesting and reproductive characteristics of coots Fulica atra breeding on two lakes in Algeria. Ardeola, 46(2): 179–186.
|
|
Ryan MR, Dinsmore JJ. 1979. A quantitative study of the behavior of breeding American Coots. Auk, 96: 704–713.
|
|
Wright A, Taylor K. 2000. Status of common loons on Squam Lake in 1999. Bird Observ, 28: 148–153.
|
|
Zhang WW, Ma JZ. 2011. Conspecific brood parasitism and defense mechanism of Fulica atra. Chinese J Zool, 46(5): 54–58.
|
Figures(7) / Tables(1)





| Study site | Code of breeding pair | Length of observation (h) | Number of platforms | Breeding period |
| Anbanghe | A | 94 | 3 | Nest building |
| Anbanghe | B | 94 | 2 | Nest building |
| Anbanghe | C | 93 | 3 | Nest building |
| Anbanghe | D | 92 | 5 | Nest building |
| Anbanghe | E* | 90 | 4 | Egg laying |
| Longfeng | F | 93 | 4 | Egg laying |
| Longfeng | G | 60 | 7 | Incubation |
| Longfeng | H | 58 | 3 | Incubation |
| Longfeng | I | 58 | 2 | Incubation |
| Longfeng | J* | 40 | ≥2 | Incubation |
| Longfeng | K* | 40 | ≥2 | Incubation |
| * We could not determine all activities of E, J and K because only part of their platforms could be observed. | ||||
DownLoad:
CSV
 Email Alerts
Email Alerts RSS Feeds
RSS Feeds