Nan WANG, Guangmei ZHENG, Philip J. K. MCGOWAN. 2012: Pheasants in sacred and other forests in western Sichuan: their cultural conservation. Avian Research, 3(1): 33-46. DOI: 10.5122/cbirds.2012.0003
Citation:
Nan WANG, Guangmei ZHENG, Philip J. K. MCGOWAN. 2012: Pheasants in sacred and other forests in western Sichuan: their cultural conservation. Avian Research, 3(1): 33-46. DOI: 10.5122/cbirds.2012.0003
Nan WANG, Guangmei ZHENG, Philip J. K. MCGOWAN. 2012: Pheasants in sacred and other forests in western Sichuan: their cultural conservation. Avian Research, 3(1): 33-46. DOI: 10.5122/cbirds.2012.0003
Citation:
Nan WANG, Guangmei ZHENG, Philip J. K. MCGOWAN. 2012: Pheasants in sacred and other forests in western Sichuan: their cultural conservation. Avian Research, 3(1): 33-46. DOI: 10.5122/cbirds.2012.0003
College of Nature Conservation, Beijing Forestry University, Beijing 100083, China
2.
Key Laboratory for Biodiversity Sciences and Ecological Engineering, Ministry of Education, College of Life Science, Beijing Normal University, Beijing 100875, China
3.
World Pheasant Association, Newcastle University Biology Field Station, Close House Estate, Heddon on the Wall, Newcastle upon Tyne, NE15 0HT, UK
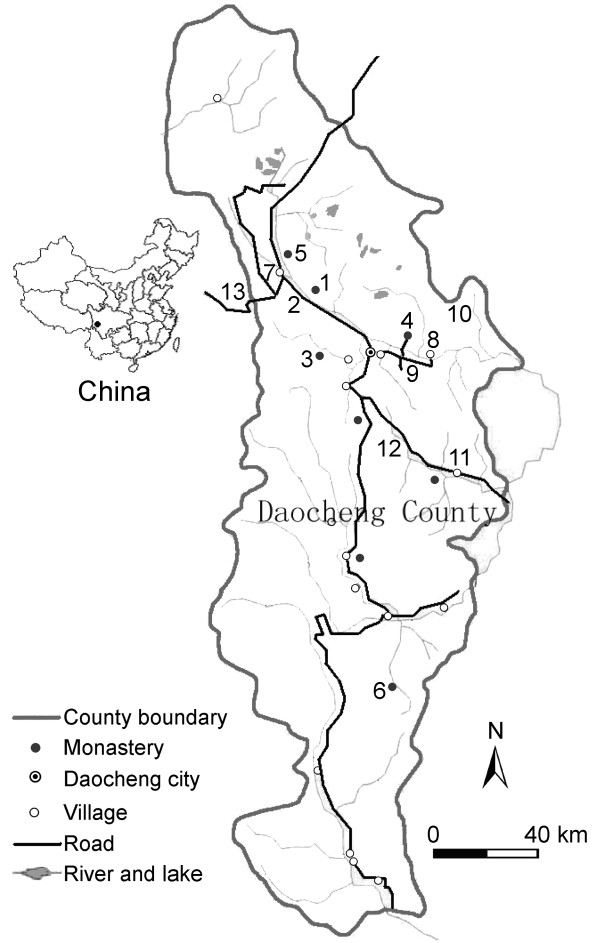
Buddhism and local cultural traditions have long protected wildlife species and their habitats in Tibetan-dominated areas of western Sichuan. In Daocheng County, the White Earedpheasant (Crossoptilon crossoptilon) has been afforded special protection by local people because it is conspicuous and white, a color with special symbolism for Buddhists. This and other cultural reasons have led to pheasants and forests benefiting in some areas. Pheasants were found during surveys between January 2003 and June 2004 in forests with varying degrees of local (non-formal) protection. However, there were significant signs that these traditional attitudes were changing in the face of three particular pressures brought to bear by better roads, improving access to and from the rest of China. The first was the development of a significant local demand for the Chinese caterpillar fungus (Cordyceps sinesis), which is much sought after throughout East Asia and mushrooms. Second, and more recent, is a dramatic increase in tourism from major Chinese cities, bringing non-Tibetan values into Daocheng County and changing the local attitudes to all animals. And then, there is a rise in income of the local population, resulting in a higher timber demand for building big houses, which impact all wildlife in the forest, but local attitudes to sacred forests have been retained so far in spite of this increased timber demand. Lessons should be learnt from the impact that unregulated tourism at Chonggu monastery, the most visited area in the county, has on the surrounding forests so that other sacred and non-sacred forests can best be protected for the long term. The alternative is that several Tibetan Plateau Galliformes, currently considered non-threatened because of their extensive distribution in a remote area, cannot be guaranteed such a healthy future.
Mothers have a variety of ways to manipulate the composition and/or environment of their eggs, including, but not limited to: hatching asynchrony (Clark and Wilson 1981; Magrath 1990), yolk hormones (Groothuis et al. 2005), sex allocation (Pike and Petrie 2003), egg size (Krist 2011), incubation temperature (DuRant et al. 2009, 2011), and carotenoids (Royle et al. 2001; Blount et al. 2002). Assessing how these factors influence the growth and survival of their offspring, however, first requires being able to accurately match hatchlings to their respective eggs. This can be a challenging endeavor for many bird species.
Frequent nest checks can be employed to directly observe the individual hatching of eggs, although increased disturbance at the nest may lead to higher rates of abandonment or predation. Even the most frequent nest checks, however, can result in finding two hatchlings within the same nest. Because egg and hatchling mass are significantly correlated in most bird species (Williams 1994; Deeming and Birchard 2007), simply assigning the heaviest hatchling to the heaviest egg (i.e., relative mass assignment; RMA) in these situations could be tempting, as RMA would increase sample sizes and reduce the number of nest checks required.
Here I present RMA as a new technique, at least to my knowledge, that may be useful in studies of maternal effects, and determine the accuracy of RMA by applying the method retrospectively to a dataset where Common Grackle (Quiscalus quiscula) hatchlings were assigned to eggs unambiguously. Common Grackles are an appropriate species in which to test the utility of RMA because hatchling mass is correlated with egg mass (Howe 1976), egg size is variable both within and among clutches (Howe 1976; Maddox and Weatherhead 2008), and 2‒4 eggs within a given clutch can hatch at approximately the same time (Howe 1976; Maddox and Weatherhead 2008).
Methods
I used an incubator with individual compartments to unambiguously assign hatchlings to their respective eggs as part of a larger study investigating maternal effects in Common Grackles. Detailed descriptions of the study site and field methods can be found in Maddox and Weatherhead(2008, 2009, 2012). Briefly, I found nests under construction and monitored them daily until fledging. On the day each egg was laid, I measured its mass (±0.1 g) and numbered it sequentially with a nontoxic felt-tip marker. When I observed at least one egg pipping from an individual nest during my daily nest checks, I collected all the eggs from the clutch and placed each egg in an individual compartment within the incubator. In total, eggs from 133 nests were treated as such. Artificial eggs of similar size and appearance were placed in nests to limit female abandonment. Upon hatching I measured the mass (±0.1 g), wing chord (±0.1 mm) and tarsus (±0.1 mm) of each hatchling before returning it to its respective nest.
When more than one egg hatched between checks of the incubator or approximately every 1‒2 h, I used RMA to assign hatchlings to their respective eggs by rank-ordering the mass of hatchlings and their candidate eggs separately and assigning hatchlings to eggs of the same rank (i.e., 1-1, 2-2, etc.). Because I knew the source egg in each case, I could evaluate the error rate of RMA. Typically two eggs from a given clutch hatched between checks but finding 3‒4 hatchlings was not uncommon. This is essentially identical to what occurs in the field with frequent nest checks.
Results
In 78 of the 133 clutches examined, two eggs hatched between consecutive checks of the incubator. RMA correctly matched 122 (78.2%) hatchlings to their respective eggs, whereas 14 (9.0%) hatchlings were incorrectly assigned. RMA could not be applied to 20 (12.8%) hatchlings because the mass of the two hatchlings or the two candidate eggs was identical. For those egg-hatchling dyads that were assigned correctly, paired differences of egg and hatchling masses averaged 0.40 g ± 0.03 SE (range 0.1-0.9 g, n = 61) and 0.38 g ± 0.04 SE (range 0.1-1.4 g, n = 61), respectively. For the 14 egg-hatchling dyads that were assigned incorrectly, paired average differences of egg (0.23 g ± 0.07 SE; range 0.1-0.6 g) and hatchling (0.21 g ± 0.04 SE; range 0.1-0.4 g) masses were almost half that of correctly assigned dyads. Excluding those hatchlings that could not be assigned, the error rate of RMA when applied to two hatchlings was 10.3% (14/136).
In 50 of the 133 clutches, three eggs hatched between consecutive incubator checks. RMA correctly matched 66 (44%) hatchlings to their respective eggs, whereas 5 clutches (10%) had at least one incorrectly assigned hatchling. Twenty-three (46%) nests contained at least one hatchling that could not be assigned to an egg for the reasons indicated above. Differences in egg mass of correct assignments averaged 0.25 ± 0.03 SE, whereas wrong assignments averaged 0.26 ± 0.02 SE. Excluding unassigned hatchlings, the error rate of RMA was 7.0% (5/71).
In the remaining nine clutches, 4 eggs within the same nest hatched between incubator checks. In one (14%) clutch, all 4 hatchlings were assigned correctly to their eggs. In another (14%) clutch, at least one hatchling was incorrectly assigned. In 7 (78%) clutches, at least one hatchling could not be assigned to an egg for the reasons indicated above. Excluding unassigned hatchlings, the error rate of RMA was 50% (1/2).
Discussion
RMA was largely successful at correctly assigning hatchlings to their respective eggs. The ability of RMA to assign eggs, however, was highly dependent on the number of hatchlings present between checks. When applied to ≥3 unknown egg-hatchling dyads the percentage of hatchlings that RMA was unable to assign was substantial, essentially eliminating its use in those situations. Consequently, I focus the remaining discussion to those cases where only two hatchlings were found between checks. With only two hatchlings present, RMA may prove to be a useful technique in some species, but its widespread adoption will likely be restricted by several shortcomings.
The accuracy of RMA is likely to vary directly with the extent that egg and hatchling mass are correlated. RMA will almost certainly perform below acceptable levels when egg mass explains little of the variation in hatchling mass. Indeed, those dyads that were incorrectly assigned generally had greater residuals than correctly assigned dyads (Fig. 1). Given that egg mass explained 82% of the variation in hatchling mass in Common Grackles—a high value for most bird species (Williams 1994)—and yet still incurred a 10% error rate, RMA should be limited to species in which egg and hatchling mass are highly correlated.
Figure
1.
Relative mass assignment performed better when the difference in egg mass was greater with incorrectly assigned eggs generally having greater residuals than correctly assigned dyads. Colors indicate correct (green), incorrect (red), or unknown (blue) assignments of egg-hatchling dyads
Restricting the application of RMA by a pre-established minimum difference in egg or hatchling mass may improve the error rate, because the pair-wise difference in mass was smaller for incorrectly assigned egg-hatchling dyads than those that were correctly assigned (Fig. 1). Limiting RMA in the current dataset to only eggs that differed in mass by > 0.1 g, reduced the error rate in half to 5%, but doubled the percentage of hatchlings that were unable to be assigned from 13 to 27%. Limiting the assignment of hatchlings had no discernible effect.
A potential pitfall of RMA not accounted for in the present study is the ability of parents to feed hatchlings between nest checks. Given the small difference in mass between nest mates, any amount of food given differentially to hatchlings could alter the mass difference relative to their nest mate and thus potentially result in a wrong assignment. This may prove negligible in many species, especially in species where males provide little parental care, as it is likely that females would still be brooding hatchlings and/or incubating eggs. Common Grackle hatchlings, however, started begging for food immediately after hatching, and I did occasionally observe males feeding hatchlings. Consequently, the error rate I report here is likely a best-case scenario.
One modification to RMA that may improve its accuracy that the current study was unable to assess is to measure egg mass directly before hatching. Given that egg mass decreases during incubation due to water loss as a result of embryonic development (Ar and Rahn 1980), egg mass obtained after this mass loss in addition to or instead of fresh egg mass may decrease the number of incorrect assignments. Future studies will need to verify if this modification would markedly increase the accuracy of RMA.
Conclusions
RMA may prove to be a valuable tool for studies that require known egg-hatchling dyads, although its accuracy should first be tested on the species in which it will ultimately be applied. Its successful application will likely be limited to those species where egg and hatchling mass are highly correlated or within-clutch egg-size variation is large. Researchers should be cognizant of the fact that RMA is a one-tailed error—egg-size effects will always be inflated—and thus could potentially result in erroneously reporting significant effects when in fact they do not exist.
Acknowledgements
I am indebted to the Shoemaker family for allowing me to work on their land and to the many people who contributed to this project over the years, especially K. Kopatz, C. Johnson, D. Kikuchi, and P. Wesley. Funding was provided by the University of Illinois, Program in Ecology, Evolution and Conservation, and the American Ornithologists' Union Research Award. L.S. Johnson provided useful comments on an earlier version of the manuscript. I was supported by an NSF International Fellowship OISE-1159178 while preparing an earlier version of this manuscript.
Competing interests
The author declares that he has no competing interests.
Ethical standards
All work described here was approved by the Institutional Animal Care and Use Committee (IACUC) at the University of Illinois (protocol #05006).
Acknowledgements
This study was supported by the National Science Foundation of China (Grant No. 30800101), the Chicago Zoological Society, the World Pheasant Association and the Critical Ecosystem Partnership Fund. Peter Garson (Chair, WPA-IUCN Pheasant Specialist Group) provided constructive advice on the work and helped facilitate financial support. We thank Alexander Pack-Blumenau and Karl-Heinz Grabowski for their assistance during the first period of fieldwork in Daocheng County and their advice. We especially thank Maotianxue for introducing us to the local people. We thank the Forestry Bureau of Sichuan Province, the Daocheng County Forestry Bureau and the Daocheng County Religious Bureau for support in various ways throughout the project.
Alemmeren Jamir S, Pandey H N. 2003. Vascular plant diversity in the sacred groves of Jaintia Hills in northeast India. Biodivers Conserv, 12: 1497–1510.
Alexander PK, Wang N, Karl-Heinz G. 2003. White-eared pheasant, monasteries and tourist development. Tragopan, 18: 18–20.
Ashalata DK, KHAN1 ML, Tripathi RS. 2005. Sacred groves of Manipur, northeast India: biodiversity value, status and strategies for their conservation. Biodivers Conserv, 14: 1541–1582.
Cheng TH. 1978. Fauna Sinica Series Vertebrata, Aves Vol. 4: Galliformes. Science Press, Beijing. (in Chinese)
Danica MA, Jan S, Robert KM, Ou X. 2005. Conserving the sacred medicine mountains: a vegetation analysis of Tibetan sacred sites in Northwest Yunnan. Biodivers Conserv, 14: 3065–3091.
Daocheng County Government. 2004. Daocheng County Government Annals. Daocheng County Government, Daocheng, Sichuan, China. (in Chinese)
Ding CQ, Zheng GM. 2000. The regions of conservation importance for grouse, partridges and pheasants in China. Bird Conserv Int, 10: 319–330.
Fuller RA, Garson PJ. 2000. Pheasants: Status Survey and Conservation Action Plan 2000–2004. IUCN, Gland, Switzerland and Cambridge, UK and World Pheasant Association, Reading, UK.
Gema JC, Dong DF, Long WX. 1999. A preliminary study of the ecology of white eared-pheasant. Chinese J Zool. 34: 26–28. (in Chinese)
Guo HC, Gao JQ, Xi QY, Li XH. 2003. Research and development for Cordyceps sinensis. J Microbiol, 23: 50–55. (in Chinese)
Huang RX, Ma L, Shao HG, Jiang T. 1990. Preliminary study on the ecology and biology of the Himalayan snowcock in Mt. Tianshan, Xinjiang. J Xinjiang Univ, 7(3): 71–75. (in Chinese)
IUCN. 2011. IUCN red list of threatened species IUCN, Gland, Switzerland and Cambridge, UK. (version 2011.2)
Jan S, Anthony A, Danica A, Kurt H, Bee G, Fang Z. 2007. Tibetan sacred sites conserve old growth trees and cover in the eastern Himalayas. Biodivers Conserv, 16: 693–706.
Jia F, Wang N, Zheng GM. 2004. Experiential model for the winter habitat selection of White Eared-pheasant flocks. J Beijing Normal Univ (Nat Sci Ed), 40: 523–530. (in Chinese with English abstract)
Jia F, Wang N, Zheng GM. 2005a. A comparison on winter habitat selection of white eared-pheasant Crossoptilon Crossoptilon and blood pheasant Ithaginis cruentus in southwest China. Bird Conserv Int, 15: 303–312.
Jia F, Wang N, Zheng GM. 2005b. Analysis of wintering roosting-site characteristics of white eared-pheasant flocks. Chinese J Ecol, 25: 153–158. (in Chinese with English abstract)
Jia F, Wang N, Zheng GM. 2005c. Habitat selection and spatial distribution of white eared-pheasant Crossoptilon crossoptilon during early breeding period. Acta Zool Sinica, 51: 383–392. (in Chinese with English abstract)
Jiang ZH. 1979. A preliminary observation of the Tibetan eared-pheasant in Tibet. Chinese J Zool, 3: 14–16. (in Chinese)
Li DH, Zheng SW, Zheng ZW. 1965. Survey of avian fauna of Yushu Qinghai. Acta Zool Sinica, 17: 217–228. (in Chinese)
Li GH, Liu LC, Zhang DY, Zhang QM. 1976. The avian fauna of Baoxing Sichuan. Acta Zool Sinica, 22: 101–114. (in Chinese)
Li T, Jia CX, Jiang YX, Sun YH. 2005. Spring habitat selection of the Chinese Grouse at Lianhuashan. Chinese J Zool, 40: 29–33. (in Chinese with English abstract)
Liu NF, Wang XT. 1990. Study of breeding ecology of Himalayan snowcock. Zool Res, 11: 299–302. (in Chinese)
Lu TC. 1991. The Rare and Endangered Gamebirds in China. Fujian Science and Technology Press, Fuzhou.
Ma M, Zhou YH, Ma L. 1991. Distribution and ecology of Himalayan snowcock. Chinese Wildl, 12(4): 15–16. (in Chinese)
Ma SL. 2004. Studies on protecting the biological diversity in Qing-Zang Plateau. J Qinghai Nation Inst (Soc Sci), 30: 76–78. (in Chinese)
Nan WY. 2001. Notion and function on nature worship of ancient Tibetan nationality. Nation Res Qinghai (Soc Sci), 12(2): 23–31. (in Chinese)
Rock JF. 1931. Konka Risumgongba, holy mountain of the outlaws. Nation Geogr M, 1931(July): 1–65.
Sang C. 2003. Tibetan traditional ecological viewpoint & ecological protection and construction. J Central Univ Natl (Phil Soc Sci Ed), 30: 13–16. (in Chinese)
Sun YH. 1997. Notes on the natural history and behaviour of the Chinese grouse Bonasa sewerzowi. Wildl Biol, 3: 265–268.
Sun YH. 2000. Distribution and status of the Chinese grouse Bonasa sewerzowi. Wildl Biol, 6: 275–279.
Tang CZ, Xu YG, Yang L. 1996. The Birds of Hengduan Mountains. Science Press, Beijing. (in Chinese)
Wang GH, Liang YF, Tian YH, Wang JL, Zhou GL. 2004. Effect of chemical insecticides on the diversity of arthropods in tea garden. J Tea Sci, 24: 99–104. (in Chinese with English abstract)
Wang N, Jia F, Zheng GM. 2005a. Comparison of the male and female behaviors of white eared-pheasant Crossoptilon crossoptilon. J Beijing Normal Univ (Nat Sci Ed), 41: 513–516. (in Chinese)
Wang N, Jia F, Zheng GM. 2005b. Nest-site selection of white eared-pheasant in southwestern Sichuan Province, China. Front Biol China, 1: 66–70.
Wang N. 2005. Winter Habitat Use and Breeding Ecology of White Eared-Pheasant Crossoptilon crossoptilon. PhD thesis. Beijing Normal University, Beijing. (in Chinese)
Xiama Z. 2002. Buddhist influence on Tibetan life, preferences for numbers and colour. China Tibet, 2002(94): 51–52. (in Chinese)
Zhang ZW, Ding CQ, Ding P, Zheng GM. 2003. The current status and a conservation strategy for species of Galliformes in China. Biodivers Sci, 11: 414–421. (in Chinese with English abstract)
Zheng GM, Wang QS. 1998. China Red Data Book of Endangered Animals: Aves. Science Press, Beijing, Hong Kong, New York. (in Chinese)
Zheng GM. 2002. A Checklist on the Classification and Distribution of the Birds of the World. Science Press, Beijing. (in Chinese)
Zheng GM. 2011. A Checklist on the Classification and Distribution of the Birds of China (Second Edition). Science Press, Beijing. (in Chinese)
Zheng SW, Pi NL. 1979. Preliminary study of Himalayan snowcock. Chinese J Zool, 14: 24–29. (in Chinese)


 DownLoad:
DownLoad:





 Email Alerts
Email Alerts RSS Feeds
RSS Feeds