In order to assess the effect of the observer distance on the behavior of Red-crowned Cranes (Grus japonensis), we studied their time budgets with a gradient of observer distances in Yancheng, China from December 2008 to March 2009. Observer distance was classified into four levels, i.e., < 200 m, 200–300 m, 300–400 m and > 400 m. The results indicated that cranes increased their vigilance when the observer was less than 300 m away. A safe distance for the observer was often as far as or more than 300 m. Cranes increased scan frequency when observers were within 200 m, whereas they prolonged the duration of their scan when observers were between 200 m and 300 m away, indicating two distinct vigilant strategies, given the observer distance. We suggest an observer distance of at least 300 m, in order to minimize the observer effect.
The presence of human observers in field behavioral studies may have a profound effect on their subjects, which is called the "observer effect" (Martin and Bateson, 1993; Lehner, 1996). In some cases, animals may respond with alarm to the observer, or escape or avoid the observer, while in other cases, animals may modify their behavior, especially their vigilance, even when they still keep doing there (MacFarlane and King, 2002). Increasing the distance between objects and observers has been reported to lessen the observer effect on behavioral time budgets in some studies (Nobbs and Mcguinness, 1999), so finding a suitable observer distance for a particular animal species can increase the accuracy of behavioral studies.
Many behavioral studies have been performed on cranes; for example, the behavior of Sandhill Cranes (Grus canadensis) (Tacha, 1988), Common Cranes (G. grus) (Aviles, 2003; Yang et al., 2006; Aviles and Bednekoff, 2007), Black-necked Cranes (G. nigricollis) (Cangjue et al., 2007; Yang et al., 2007) and Red-crowned Cranes (G. japonensis) (Liu et al., 2001; Zhou et al., 2002; Zou et al., 2003). Most of these studies are based on observers being present in the field, observing and recording behavior, but no one reported the distance between the observer and cranes. Although some researchers report cranes usually fly away when observers approach within 100 m (Ma et al., 2000), no detailed study of observer distance on the time budgets has been carried out. For our study, we have used Red-crowned Cranes, endemic to east Asia, to study the effect of observer distance on the time budgets of these cranes.
Materials and methods
Study area
This study was conducted in the buffer zone of the Reed Company of the Yancheng Biosphere Reserve (32°34′–34°28′N, 119°48′–120°56′E) in Jiangsu Province, China. The reserve was established in 1983 for conservation of Red-crowned Cranes and other waterbirds and was internationally recognized as a biosphere reserve by UNESCO in 1992. The coastline of the reserve is about 580 km long and the area of the reserve is about 2400 km2. Mean temperature in winter is about 4℃ and varies from –8℃ to 16℃. Snow covers the ground occasionally in winter. The dominant plants are Phragmites australis, Suaeda salsa and Imperata cylindrica.
The Red-crowned Crane, which is listed in the Convention on Migratory Species (CMS) Appendix 1 and CITES Appendix 1, winters along rivers and coastal marshes in north-eastern Asia (BirdLife-International, 2011). A large proportion of the migratory population of Red-crowned Cranes winters in the Yancheng Biosphere Reserve. Red-crowned Cranes migrate from north-eastern China to Yancheng in late October and overwinter in the reserve until early March. The wintering crane population numbered more than 1000 birds in 2001 (Ma et al., 2009), but fewer than 500 cranes remained in the 2010 winter survey.
Observations on behavior
Wintering cranes are gregarious in small family or large social groups (Ma et al., 2000; Wang et al., 2011). A family group is composed of two adult cranes, with 0–2 juvenile cranes, while a social group is made up of a number of family groups. We selected family groups as our targets, in order to remove the effect of group size on individual behavior. Observations of their behavior were conducted from late December 2008 to early March 2009. Cranes were located during regular circuits with a GPS and a rangefinder. The circuit was not repeated on the same day in order to avoid replicated sampling. Observations were not made on days with rain, snow or strong winds to lessen bias caused by the effect of extreme weather.
Focal samplings were carried out using binoculars (8×56) or a telescope (20–60×63) (Martin and Bateson, 1993). Observer distances were determined whenever the observer could record the exact behavior of the cranes. Then distance was estimated with a rangefinder and classified into four levels: i.e., < 200 m, 200–300 m, 300–400 m and > 400 m. We also recorded the date, time of day, location, habitat (farmland, reed beds, grassland), group size (including juveniles), group composition (with or without juveniles) and the number of juveniles. Focal individuals were randomly selected from a family group. At most we selected one adult and one juvenile from the same group to avoid resampling.
Behavioral events were dictated on to a MP3 recorder. Observations lasted 30 min unless we lost sight of the individual bird in focus. Actual observation time ranged from 10 min to 30 min, with an average of 24 min. We distinguished five behavioral states: feeding, vigilance, preening, locomotion and other behavior. Feeding refers to a crane excavating food, swallowing or drinking. Vigilance refers to a crane stretching its head upwards while looking around. Preening includes combing feathers, shaking head, or stretching legs. Locomotion includes walking, running, leaping and flying. Other behavior includes resting, fighting and calling.
Data analysis
Because feeding and vigilance made up more than 90% of the total time budgets, we only analyzed these two behavior categories. From the timed sequence of events, time spent feeding and in vigilance was summed respectively across focal observations and expressed as a percentage of total observation time. Frequencies for feeding, vigilance and total behavioral bouts were also counted as number per min. For our analysis, percentages of time spent feeding and in vigilance were changed by an arcsine square-root transformation. Frequencies for feeding, vigilance and total behavioral bouts were normally distributed given the one-sample K-S test we used (p > 0.905). In our model, the following four variables were compared with a general linear model (GLM): time of day (8:00–11:00, 11:00–14:00, 14:00–18:00), habitat (reed beds, farmland), age (adult, juvenile) and distance (< 200 m, 200–300 m, 300–400 m, > 400 m). No significant differences were found for time, habitat and their interactions (p > 0.05), so we removed these factors from the model. Therefore, the final model only included the factors age and distance; post-hoc testing was carried out with S-N-K tests. All analyses were carried out with SPSS 13.0. The level of statistical significance was set at 0.05 throughout. We reported mean and standard errors obtained from the statistical models.
Results
We obtained 126 focal observations representing a total of 2926 min of observations. A total of 70 focal observations (56%) originated from adults while 56 samples (44%) were obtained from juveniles. There were, respectively, 30, 33, 27 and 36 samples for distances < 200 m, 200–300 m, 300–400 m and > 400 m.
On average, cranes spent 64% and 27% of their time in feeding and vigilance, while the other types of behavior accounting for only about 9%. The average frequency for feeding, vigilance and behavior transition was 0.808, 0.851 and 1.945 times per min respectively.
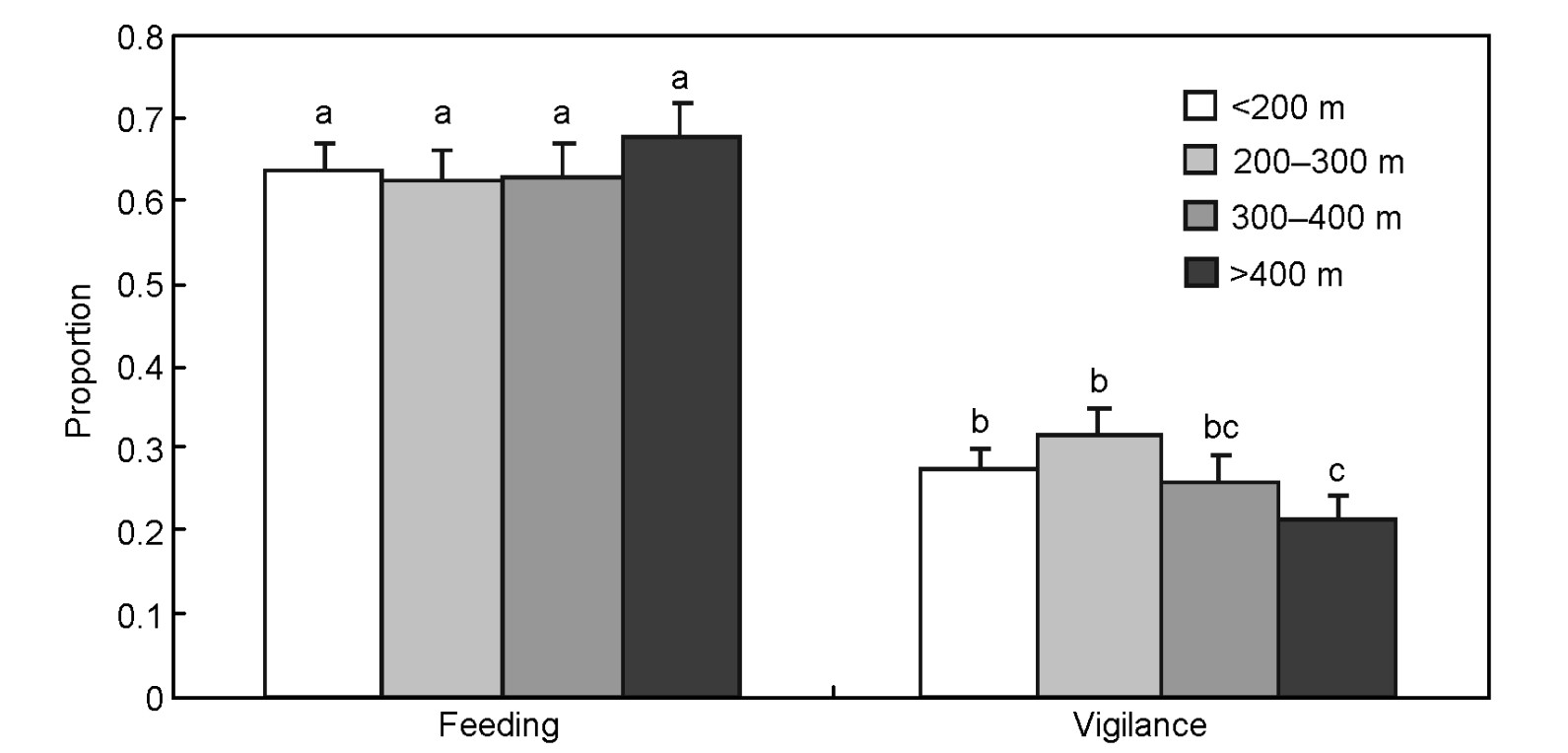
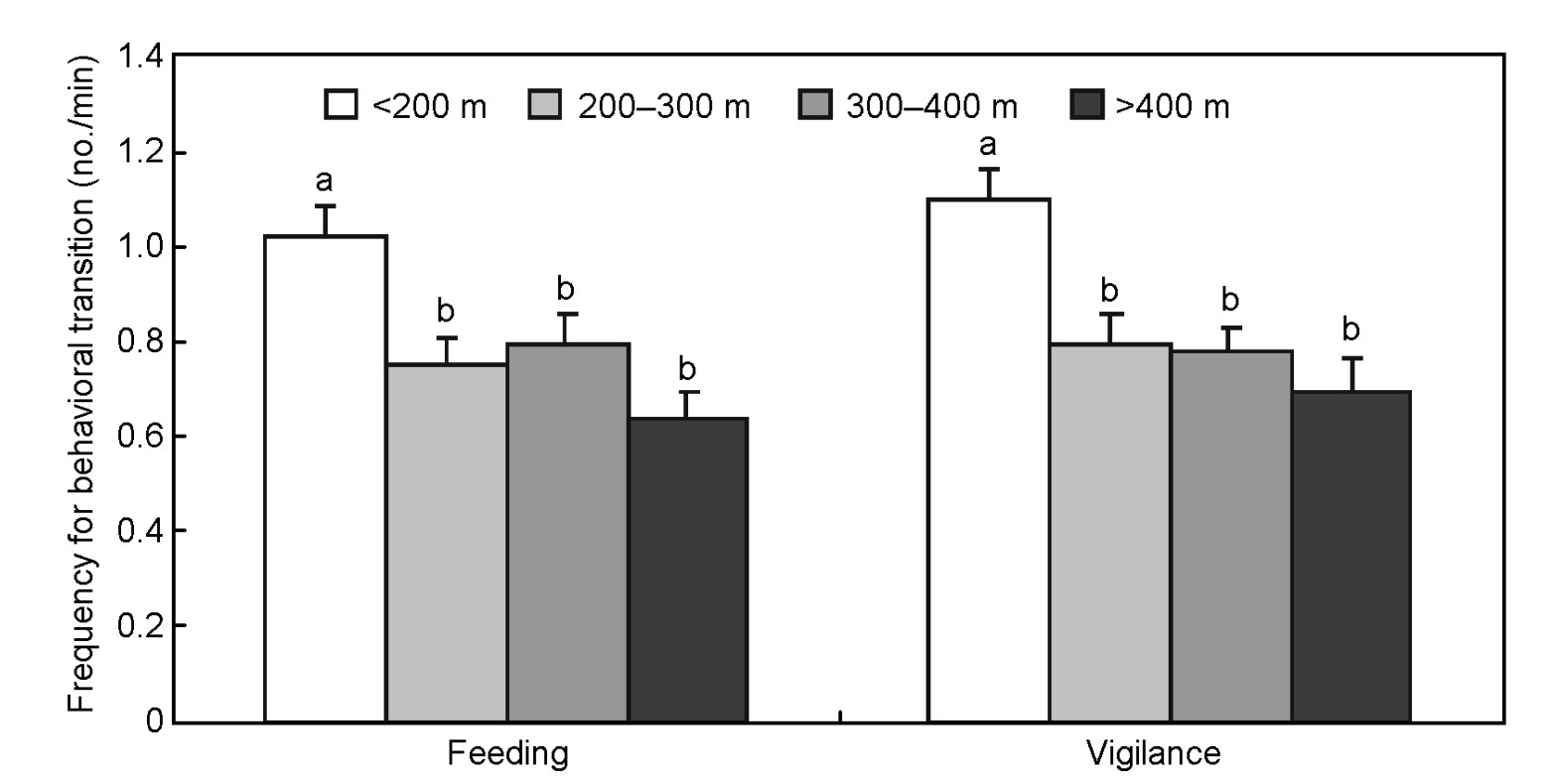
The time budget of cranes in vigilance (F3,121 = 5.259, p = 0.002, Fig. 1), frequencies in feeding (F3,121 = 6.450, p < 0.001), frequencies in vigilance (F3,121 = 9.645, p < 0.001) and total behavioral bouts (F3,121 = 9.844, p < 0.001, Fig. 2) were dependent on the distance of the observer. Post-hoc tests indicated that cranes spent less time in vigilance (0.215 ± 0.026) when the observer was beyond 400 m distance than within 300 m (≥ 0.257 ± 0.032). Frequencies of feeding (Ff), vigilance (Fv) and total behavioral bouts (Ft) were clearly higher when the observer was within 200 m distance (Ff = 1.023 ± 0.062, Fv = 1.097 ± 0.065, Ft = 2.478 ± 0.121) than beyond 200 m (Ff ≤ 0.790 ± 0.061, Fv ≤ 0.794 ± 0.059, Ft ≤ 1.867 ± 0.113). No difference in time budgets of feeding (F3,121 = 1.153, p = 0.331) was found between distances.
Figure
1.
Time budgets of red-crowned cranes along a gradient of observer distance. Same letters upon the strips denote no difference (p > 0.05).
The presence of an observer may represent a threat stimulus, or a risk of predation, which may in turn increase defensive behavior, such as vigilance (Frid and Dill, 2002; MacFarlane and King, 2002; de Almeida et al., 2006). The shorter the distance between the observer and cranes, the greater the potential of threat. Thus a greater proportion in vigilant behavior would be expected to appear within shorter distances. Moreover, the increase in vigilant behavior might also be reflected as an incremental scan frequency or a prolonged scan duration (Beauchamp, 2008), where the former would lead to a faster frequency in the number of total behavioral bouts.
We did, in fact, find a decrease in the proportion of vigilant behavior with an increase in observer distance. Cranes spent more time in vigilance when the observer was within 300 m, whereas they behaved least vigilantly when the observer was farther than 400 m. Although a previous study reported that cranes would fly away when observers were within 100 m (Ma et al., 2000), our results indicated that cranes actually modified their vigilance even when observers were within 300 m. The safe distance or so-called vigilant distance for Red-crowned Cranes would be as far as or more than 300 m. This vigilant distance is affected by a number of factors, such as body size, habitat and others (Fernandez-Juricic et al., 2001; Fernandez-Juricic et al., 2002). However, all 15 crane species are large waders, most with body length more than 100 cm and most of them live in open wetlands, such as tidal grasslands (Ellis et al., 1996), so we consider that a safe observer distance of 300 m for Red-crowned Cranes may also be effective in other cranes. We did not find a significant difference in feeding budget, suggesting that the observer effect was absent.
The frequency of behavioral bouts, including feeding and vigilance, were far greater within 200 m than beyond this distance. Similar to other cranes (Tacha, 1988; Aviles, 2003; Cangjue et al., 2007), feeding and vigilance were the two main types of behavior of the Red-crowned Cranes, which accounted for about 90% of their total time budgets. When observers were within 200 m, cranes clearly increased their scan frequency, which caused a similar increase in feeding frequency. When the distance was more than 200 m, there was no difference in behavioral transition frequency. We have known that a safe distance for cranes is about 300 m, but we found that the frequency of their behavior did not change between 200 m and 300 m from that within 200 m, i.e., cranes prolonged their scan duration when observers were between 200 m and 300 m (χ2 = 7.063, df = 1, p = 0.018). Different observer distances might represent different levels of predation risk. Cranes clearly adopted two distinct strategies to increase the time of vigilance according to different observer distances. They increased their scan frequencies when the distance was less than 200 m, but prolonged the time for scanning when it was between 200 m and 300 m.
We conclude that the observer effect has an obvious impact upon the behavior of Red-crowned Cranes. The presence of an observer increases vigilance significantly within 300 m. Cranes respond to this presence by increasing their scan frequency when the observer was less than 200 m away and prolonged it when the distance was between 200 m and 300 m. Based on our results, we suggest investigators of cranes should observe their targets from at least a 300 m distance, in order to minimize the observer effect.
Acknowlegements
We received financial support from the Chinese Key Technology R & D Program of the Eleventh Five-year Plan (No. 2008BAC39B03) and the Nanjing University Funds for Starting Research. We thank Dr. Liying SU, ICF, for her kind suggestions about this study. We also thank Jing Li, Chen Ge, Zhi Wang, Guorong Sun, Zhijun Song, Zhifei Song and Luping Zhang for their help during field work.
Aviles JM, Bednekoff PA. 2007. How do vigilance and feeding by common cranes Grus grus depend on age, habitat, and flock size? J Avian Biol, 38: 690–697.
Aviles JM. 2003. Time budget and habitat use of the Common Crane wintering in dehesas of southwestern Spain. Can J Zool, 81: 1233–1238.
Beauchamp G. 2008. What is the magnitude of the group-size effect on vigilance? Behav Ecol, 19: 1361–1368.
BirdLife International. 2011. Species factsheet: Grus japonensis. . Accessed on 18 March 2011.
Cangjue Z, Yang L, Li J. 2007. Time Budget in daytime of Black-necked Crane in wintering period in Tibet. Chinese J Wildlife, 29: 15–20. (in Chinese with English abstract)
de Almeida M, de Paula H, Tavora E. 2006. Observer effects on the behavior of non-habituated wild living marmosets (Callithrix jacchus). Rev Etol, 8: 81–87.
Ellis DH, Gee GF, Mirande CM. 1996. Cranes: Their Biology, Husbandry, and Conservation. National Biological Service / International Crane Foundation Limited Edition, Washington.
Fernandez-Juricic E, Jimenez MD, Lucas E. 2001. Alert distance as an alternative measure of bird tolerance to human disturbance: implications for park design. Environ Conserv, 28: 263–269.
Fernandez-Juricic E, Jimenez MD, Lucas E. 2002. Factors affecting intra- and inter-specific variations in the difference between alert distances and flight distances for birds in forested habitats. Can J Zool, 80: 1212–1220.
Frid A, Dill L. 2002. Human-caused disturbance stimuli as a form of predation risk. Conserv Ecol, 6: 11.
Lehner PN. 1996. Handbook of Ethological Methods. Cambridge University Press, New York.
Liu Z, Wu J, Li X, Teng L, Wang X, Guo W, Qi Z, Yao H, Qiu F. 2001. Behavior of Red-crowned Crane during breeding season in Zhalong Nature Reserve. J Northeast Forest Univ, 29: 92–95. (in Chinese with English abstract)
Ma ZJ, Li B, Li WJ, Han NY, Chen JK, Watkinson AR. 2009. Conflicts between biodiversity conservation and development in a biosphere reserve. J Appl Ecol, 46: 527–535.
Ma ZJ, Li WJ, Wang ZJ. 2000. Natural Conservation of Red-crowned Cranes. Qinghua University Press, Beijing. (in Chinese)
MacFarlane G, King S. 2002. Observer presence influences behaviour of the semaphore crab, Heloecious cordiformis. Anim Behav, 63: 1191–1194.
Martin P, Bateson P. 1993. Measuring Behaviour: An Introductory Guide. Cambridge University Press, Cambridge.
Nobbs M, Mcguinness K. 1999. Developing methods for quantifying the apparent abundance of fiddler crabs (Ocypodidae: Uca) in mangrove habitats. Austral Ecol, 24: 43–49.
Tacha T. 1988. Social organization of sandhill cranes from midcontinental North America. Wildlife Monogr, 99: 3–37.
Wang Z, Li ZQ, Beauchamp G, Jiang ZG. 2011. Flock size and human disturbance affect vigilance of endangered red-crowned cranes (Grus japonensis). Biol Conserv, 144: 101–105.
Yang R, Wu HQ, Yang XJ, Jiang WG, Zuo L, Xiang ZR. 2007. Diurnal time budget of the Black-necked Crane during the breeding season. Waterbirds, 30: 80–85.
Yang Y, Chen W, Jiang W, Yang S, Peng G, Huang T. 2006. Effects of group size on vigilance behavior of wintering common cranes. Zool Res, 27: 357–362.
Zhou X, Liu Z, Wu J, Teng L, Wang X. 2002. Time budget and activity rhythm of Red-crowned crane during breeding season. J Northeast Forest Univ, 30: 60–62. (in Chinese with English abstract)
Zou H, Wu Q, Piao L. 2003. Time budget of Red-crowned crane during Incubating Season in Zhalong Nature Reserve after burning and irrigating. J Northeast Forest Univ, 31: 41–42. (in Chinese with English abstract)

 DownLoad:
DownLoad:





 Email Alerts
Email Alerts RSS Feeds
RSS Feeds