Xin LEI, Zuohua YIN, Zhenmin LIAN, Cungen CHEN, Chuanyin DAI, Anton KRIŠTÍN, Fumin LEI. 2010: Phylogenetic relationships of some Sylviidae species based on complete mtDNA cyt b and partial COI sequence data. Avian Research, 1(3): 175-187. DOI: 10.5122/cbirds.2010.0013
Citation:
Xin LEI, Zuohua YIN, Zhenmin LIAN, Cungen CHEN, Chuanyin DAI, Anton KRIŠTÍN, Fumin LEI. 2010: Phylogenetic relationships of some Sylviidae species based on complete mtDNA cyt b and partial COI sequence data. Avian Research, 1(3): 175-187. DOI: 10.5122/cbirds.2010.0013
Xin LEI, Zuohua YIN, Zhenmin LIAN, Cungen CHEN, Chuanyin DAI, Anton KRIŠTÍN, Fumin LEI. 2010: Phylogenetic relationships of some Sylviidae species based on complete mtDNA cyt b and partial COI sequence data. Avian Research, 1(3): 175-187. DOI: 10.5122/cbirds.2010.0013
Citation:
Xin LEI, Zuohua YIN, Zhenmin LIAN, Cungen CHEN, Chuanyin DAI, Anton KRIŠTÍN, Fumin LEI. 2010: Phylogenetic relationships of some Sylviidae species based on complete mtDNA cyt b and partial COI sequence data. Avian Research, 1(3): 175-187. DOI: 10.5122/cbirds.2010.0013
Members of the passerine family Sylviidae are distributed widely around the Old World and the phylogenetic relationships of many species still remain controversial. In this study, we investigated the phylogeny and relationships among 37 species of 11 genera by analyzing DNA sequences obtained from the complete mitochondrial cytochrome b (cyt b) and partial cytochrome oxidase I (COI) genes. The data were analyzed by maximum-likelihood analysis and Bayesian inference. The results demonstrate that the current genus Phylloscopus is non-monophyletic, while Seicercus is synonymized with Phylloscopus or is a group within the genus Phylloscopus. We may conclude that within Phylloscopus there are close relations between P. magnirostris and P. borealis; among P. proregulus, P. yunnanensis, P. pulcher and P. maculipennis, as well as among P. occisinensis, P. schwarzi, P. armandii, P. collybita and P. fuscatus. Monophyly of the genus Cettia could not be corroborated; it is closely related to the genera Abroscopus and Tesia. However monophyly of Acrocephalus is supported. Furthermore, the close relationships between Sylvia and Zosterops and among Prinia, Orthotomus and Cisticola are also supported.
Sylviidae, a family of small Old World warblers, has proven to be a controversial group and, for a long time, problematic in taxonomy, owing to subtle morphological distinctions in many species and subspecies (Zheng, 2005). Traditionally, taxonomists considered that these warblers should be grouped in the subfamily Sylviinae, consisting of 60 genera and 348 species (Mayr and Cottrell, 1986). Many previous studies on the relations among the Sylviidae species and their taxonomic status are mostly based on morphological and ecological characteristics (La Touche, 1925–1934; Vaurie, 1965; Mayr and Cottrell, 1986). However, cryptic species are quite common in Sylviidae and sonogram analysis and molecular genetic approaches have been frequently used to solve issues of species delimitation and taxonomic relationships (Drovetski et al., 2004; Alström et al., 2006). Since the first analyses of DNA-DNA hybridization, Sylviinae has been elevated as the family Sylviidae and divided into four subfamilies: Acrocephalinae, Megalurinae, Garrulacinae and Sylviinae, while some genera have been modulated (Sibley and Monroe, 1990). After this, most species were studied using molecular approaches and their relationships were revised repeatedly, especially in Europe (Helbig and Seibold, 1999; Drovetski et al., 2004; Alström et al., 2006).
In China, based on traditional morphological taxonomic approaches, Cheng(1994, 2000) recognized 98 or 95 species in 18 genera and listed them in the subfamily Sylviinae under the family Muscicapidae, including the genera Tesia, Cettia, Bradypterus, Megalurus, Locustella, Acrocephalus, Hippolais, Sylvia, Phylloscopus, Regulus, Seicercus, Abroscopus, Tickellis, Leptopoecile, Orthotomus, Cisticola, Graminicola and Prinia. Recently, Cheng's classification was revised by Zheng (2005) mostly based on Sibley and Monroe's taxonomic treatment and sequences, in which Sylviinae has been elevated to the family Sylviidae consisting of 16 genera, while Regulus was promoted as the family Regulidae, while Cisticola and Prinia were placed (similar as in Dickinson (2003)) in the family Cisticolidae. Although a few studies on the phylogenetic relations of the species are available from Asia, there are still many taxonomic problems about the relationships among some genera, especially from China which harbors abundant warbler diversity and the classification and phylogenetic relationships of many more putative species and genera in Sylviidae still remain unsolved (Alström et al., 2007, 2008; Martens et al., 2008; Päckert et al., 2009). In our study we investigated the phylogenetic relationships among genera and some species of Sylviidae, based on sequence data of mitochondrial DNA, in an attempt to reconstruct a phylogenetic topology for the constituents of this group and to assess the validity of the taxonomic status of some controversial genera and species.
The mitochondrial cytochrome b (cyt b) gene is the most widely used genetic marker for phylogenetic studies and has been the most readily available source of sequence data in avian studies (Johnson, 2001; Klicka et al., 2001; Thomassen et al., 2003; Sheldon et al., 2005). Cytochrome oxidase I (COI) gene is also a very useful tool for DNA-barcoding, allowing studies of avian species delimitation and their phylogenies (DeFilippis, 1995; Weibel and Moore, 2002; Hebert et al., 2004; Webb and Moore, 2005; Aliabadian et al., 2009). In this study, we selected species as in-group following Zheng's classification of Sylviidae (Zheng, 2005) and then investigated the phylogeny and relations among some species and genera by DNA sequencing of the complete cyt b and partial COI genes.
Materials and methods
Selection of in-group taxon and out-groups
We included 36 Sylviidae species in the study (Table 1). In attempting to enhance viewing the phylogenetic relationships of Sylviidae species, we also included Zosterops japonica from Zosteropidae. We used Lanius isabellinus and Dicrurus hottentottus as out-groups. Samples were collected mostly from China. Only the Locustella fluviatilis, Sylvia communis and Phylloscopus collybita species are from Slovakia (Europe). All birds were collected complying with the current laws in China and Slovakia.
Table
1.
Species list, samples used, mitochondrial DNA cyt b and COI gene sequences
Total genomic DNA was extracted from blood or muscle specimens using the TIANamp Genomic DNA Kit (TIANGEN) as per instructions of the manufacturer. Nucleotide sequence data were obtained from the mitochondrial cyt b gene and COI.
The primers used to amplify the cyt b gene were L14827 and H16065 (Pasquet et al., 2002), L14731 and H16067 (Saetre et al., 2001), L14851 and H16058 (Groth 1998), L14863 and H16058 (Groth, 1998). The primers L6615 and H7956 (Sorenson et al., 1999) were used for the COI gene. Amplification products were sequenced with the same primers as used for PCR amplification.
PCR reactions were carried out under the following conditions: an initial denaturation at 94℃ for 8 min; 36 cycles at 94℃ for 30 s, 45–48℃ for 1 min and 72℃ for 2 min, followed by a final extension of 10 min at 72℃. For all taxa, both strands of DNA were sequenced using an ABI3730 automated sequencer. The DNA sequences are deposited at GenBank (accession number from HQ608821 to HQ608894).
Alignment and sequence properties
All DNA sequence datasets were edited using the DNASTAR package (SeqMan), and the sequences of the two gene regions were aligned using Clustal W1.83 (Thompson et al., 1997). No gaps, insertions, or deletions were found in the aligned sequences and all sequences were translated into amino acid sequences to verify the alignments. Both separated and combined datasets were analyzed. The final sequences included complete cyt b gene (1143 bp) and part of COI gene (1176 bp). Statistics for nucleotide variation and pairwise genetic distances were computed with MEGA 3.1 (Kumar et al., 2004).
Phylogenetic analyses
Phylogenetic analyses were performed on the combined sequences from the cyt b and COI genes. In addition, phylogenetic signals in the two datasets were compared by analyzing each gene region separately.
Maximum-likelihood (ML) analyses and incongruence length difference (ILD or partition homogeneity) tests were performed using Paup* 4.0b10 (Swofford, 2002). For ML, the optimal model of evolution was determined by hierarchical likelihood ratio tests (hLRTs) in Modeltest 3.06 (Posada and Crandall, 1998). Parameters for the ML analyses were estimated from the data (Table 2). Furthermore, the GTR + I + G model was identified as the best fit for our data using hLRTs criteria in Modeltest. Bootstrap support values were based on 100 replicate, maximum-likelihood analyses.
Table
2.
Observed pairwise genetic distances for the cyt b gene (below diagonal) and the COI gene (above diagonal)
The datasets were also analyzed by Bayesian inference. The models for nucleotide substitutions were selected for the two genes individually using the Akaike Information Criterion (Akaike, 1973). We ran four Markov chains for 5 million generations each with trees sampled every 100 generations. The trees saved during the "burn-in phase" (the first 100000 generations in each analysis) were discarded. The posterior probabilities were then calculated from the remaining 49000 saved trees. The remaining trees from both analyses (produced automatically in MrBayes v3.1b) were used to create a majority rule consensus tree. Posterior probabilities greater or equal to 95% were considered significant (Leache and Reeder, 2002).
Results
Sequence characteristics
In cyt b, 523 of 1143 sites varied among taxa and 448 sites (39%) were parsimony-informative. The COI gene was less variable than cyt b: 424 of 1176 sites varied among taxa and 386 sites (33%) were parsimony-informative. The combined sequences of the two gene segments had 2319 sites, of which 834 (36%) were parsimony-informative.
Pairwise distances among the 37 in-group species and 2 out-group species are summarized in Table 2. In cyt b gene, the observed intra-generic sequence divergence ranged from 0.001 (Acrocephalus orientalis and A. aedon, Phylloscopus proregulus and P. yunnanensis) to 0.138 (Phylloscopus collybita and P. trochiloides). Inter-generic cyt b comparisons ranged from 0.118 (Phylloscopus coronatus and Cettia diphone, Prinia criniger and Orthotomus sutorius) to 0.204 (Sylvia curruca and Acrocephalus aedon). The smallest divergence in cyt b between the in-group and out-group was 0.182 (Locustella lanceolata and Lanius isabellinus), and the largest 0.232 (Sylvia curruca and Dicrurus hottentottus). In the COI gene, the smallest intra-generic sequence divergence within the in-group was 0.001 (Phylloscopus proregulus and P. yunnanensis, Phylloscopus schwarzi and P. armandii, Acrocephalus orientalis and A. aedon) and the largest 0.144 (Phylloscopus collybita and P. maculipennis). Inter-generic COI comparisons ranged from 0.111 (Cettia fortipes and Abroscopus albogularis) to 0.176 (Prinia criniger and Seicercus castaniceps, Cisticola juncidis and Phylloscopus trochiloides). The smallest divergence observed between the in-group and the two out-groups was 0.150 (Orthotomus sutorius and Dicrurus hottentottus, Sylvia communis and Dicrurus hottentottus) while the largest divergence was 0.187 (Acrocephalus bistrigiceps and Dicrurus hottentottus).
cyt b and COI had very similar nucleotide compositions, so the two genes, when combined, had a more uniform nucleotide composition than any individual gene. Nucleotide bias of the two genes was similar to that observed in birds in previous studies (Weibel and Moore, 2002; Webb and Moore, 2005). At the first codon position, the four bases were equally distributed. At the second position, the amount of G was decreased and that of T increased. The strong bias for an excess of C and paucity of G was shown at the third codon positions.
Phylogenetic analysis
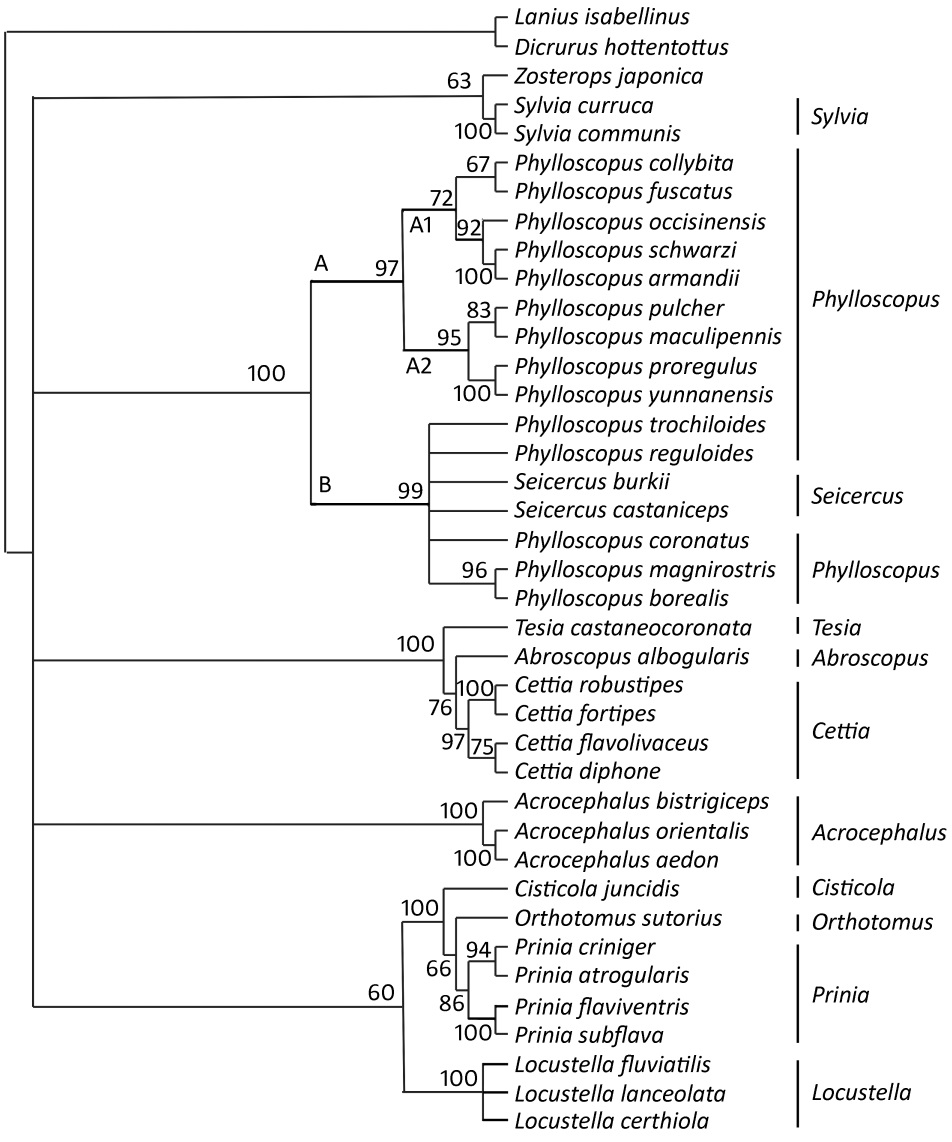
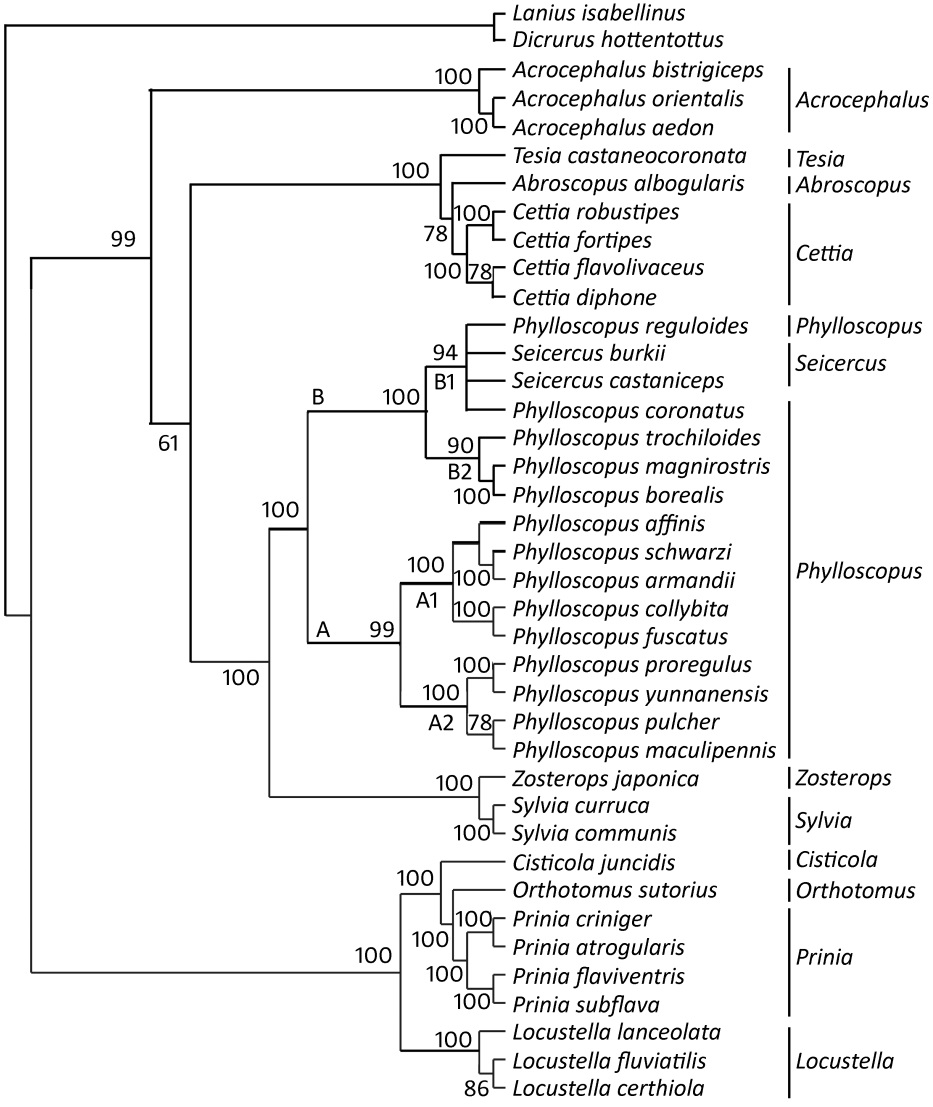
We analyzed the topologies of ML and Bayesian trees produced by the combined sequences of the two gene segments. The trees, resulting from the maximum-likelihood analysis and Bayesian inference have practically identical topologies when the frequency of occurrence is set to 50% (Figs. 1 and 2).
Figure
1.
The maximum likelihood tree (Bootstrap values are shown at nodes on the maximum likelihood trees.) from analysis of the cyt b and COI sequences
Figure
2.
The Bayesian tree (The mean posterior probabilities on the Bayesian tree are given only where they were 50% or higher) from analysis of the cyt b and COI sequences
The taxa fall into five major clades. Sylvia and Zosterops are clustered within Clade 1 (ML: 63%; Bayesian: 100%). In Clade 2, Seicercus is nested within Phylloscopus (ML: 99%; Bayesian: 94%) and the latter genus is divided into three clades in ML tree: Clade A1 with P. collybita, P. fuscatus, P. occisinensis, P. schwarzi and P. armandii (ML: 72%; Bayesian: 100%), Clade A2 with P. pulcher, P. maculipennis, P. proregulus and P. yunnanensis (ML: 95%; Bayesian: 100%) and Clade B with P. trochiloides, P. reguloides, P. coronatus, P. magnirostris, P. borealis, Seicercus burkii and S. castaniceps (ML: 99%, Bayesian: 100%). However, in the Bayesian tree, Clade B is divided into two small clades: Clade B1 with P. reguloides, P. coronatus, Seicercus burkii and S. castaniceps (94%) and Clade B2 with P. trochiloides, P. magnirostris and borealis (90%). The close relationship among Tesia, Abroscopus and Cettia receives good bootstrap and posterior probability support (ML: 100% and 76%; Bayesian: 100% and 78%). Clade 4 comprises only three members of Acrocephalus. Locustella, Prinia, Orthotomus and Cisticola are clustered within Clade 5 (ML: 60%; Bayesian: 100%). Our results here show a close relationship among Cisticola, Orthotomus and Prinia with good nodal support (ML: 100% and 66%; Bayesian: 100% and 100%).
In our study, the maximum-likelihood and Bayesian analyses both suggest that Seicercus is a close relative of Phylloscopus, especially of P. reguloides, P. coronatus, P. trochiloides, P. magnirostris and P. borealis. Although only two species of Seicercus (S. burkii and S. castaniceps) were studied, we strongly support the idea that the monophyly of Phylloscopus is invalid (Olsson et al., 2004, 2005). Two species of Seicercus were grouped with five species of Phylloscopus (Clade B) and the largest genetic distance among them (0.117) was lower than the largest distance among 14 species of Phylloscopus (0.138). Furthermore, Phylloscopus and Seicercus species have many similar morphological characters, such as incompact feathers on forehead, prolonged shaft propers, many supplementaries before rectal bristles and twelve tail feathers. In view of this evidence, we support the viewpoint that Phylloscopus is non-monophyletic, which should include Phylloscopus and Seicercus, and suggest that Phylloscopus and Seicercus could be combined into one genus and that the complete species of these two former genera are necessarily involved in further review.
Relationships within Phylloscopus
The genus Phylloscopus has the most taxonomic problems. Little is known about this genus in China, except for the morphological review by Jia et al. (2003). As well, new species in Phylloscopus have frequently been found (Olsson et al., 2005; Martens et al., 2008; Päckert et al., 2009), e.g. twelve new species were found in China over a period of ten years during the last century (Irwin et al., 2001). Therefore, a taxonomic revision of some species and subspecies is still needed. However, taxonomic arrangements have traditionally relied on similarities in morphology and ecology (Cheng, 1994, 2000; Zheng, 2005). Based on DNA sequence data from our current study with strong support from some closely related species, allow us to cast new insights into the evolution of these birds.
We found two deeply distinct divergent clades (Clades A and B) of Phylloscopus in both maximum-likelihood and Bayesian trees (Figs. 1 and 2). Clade A includes two small clades: Clade A1 (including P. collybita, P. fuscatus, P. occisinensis, P. schwarzi and P. armandii) and Clade A2 (including P. pulcher, P. maculipennis, P. proregulus and P. yunnanensis). Clade B includes five species of Phylloscopus in a ML tree and there is a close relationship between P. magnirostris and P. boreali. Clade B is divided into two sister groups in a Bayesian tree: Clade B1 (including P. reguloides, P. coronatus, Seicercus burkii and S. castaniceps) and Clade B2 (including P. trochiloides, P. magnirostris and P. borealis). The positions of these species were stable and strongly supported by the trees. Our molecular results are also corroborated by some morphological and ecological characters. There are some morphological similarities of P. magnirostris and P. boreali among these three species, except for the sixth primary remiges, for example the olive green body and a pair of brown wings. In Clade A1, species P. occisinensis, P. collybita, P. fuscatus, P. schwarzi and P. armandii share the same morphological character (no stripes on the wings). Species P. proregulus, P. pulcher, P. maculipennis and P. yunnanensis in Clade A2 have some distinct morphological characters (one yellow caestus and two yellow stripes on the wings) and inhabit elevations above 1500 m a.s.l. Olsson et al. (2005) also supported the two close relations between P. proregulus and P. maculipennis and between P. collybita and P. schwarzi on the basis of DNA analysis (cyt b, 12S and myoglobin intron II). We may conclude then that there are close relationships between P. magnirostris and P. borealis, among P. proregulus, P. yunnanensis, P. pulcher and P. maculipennis, among the following five species, P. occisinensis, P. collybita, P. fuscatus, P. schwarzi and P. armandii. Because the Clade B1 and Clade B2 were not supported on the ML trees, the relationships among these species cannot be resolved in this study.
However, there are currently over 30 Phylloscopus species recognized in China and over 50 across the world (Monroe and Sibley, 1993; Zheng, 2005). Unfortunately, because only 16 representatives from Phylloscopus and Seicercus were included in this study, the validity of Phylloscopus is premature for a revision by us and the suggestion needs to be proven in future studies.
Taxonomic status of Sylvia and Zosterops
Monroe and Sibley (1993) considered the Sylvia genus within Sylviidae and Zosterops genus in Zosteropidae (both families in the superfamily Sylvioidea). Cheng (2000) placed Sylvia into Sylviinae under the family Muscicapidae and Zosterops into Zosteropidae. Mackinnon and Phillipps (2000) and Zheng (2005) also considered that Sylvia and Zosterops fell into two separate families, Sylviidae and Zosteropidae. Recently, a close association of Zosterops and Sylvia has been suggested by several studies on the basis of mitochondrial and nuclear DNA sequences (Barker et al., 2002, 2004; Cibois, 2003; Ericson and Johansson, 2003). Furthermore, Alström et al. (2006) showed that Sylvia, Zosterops, Garrulax and Timaliini are clustered within the same clade and suggested the name Timaliidae for this clade. Although few morphological similarities exist between Zosterops and Sylvia, their close relationship is strongly supported in our study, based on mitochondrial gene sequences. However, we have only one sample of Zosterops and two samples of Sylvia. A phylogenetic study of these two genera should be considered, at best, as uncertain, but needs to be undertaken in the future.
Relationships among Cettia, Abroscopus and Tesia
The genera Cettia, Abroscopus and Tesia were placed in Acrocephalinae by Sibley and Monroe (1990). Some taxonomists considered that Cettia, Tesia and Urosphena were near relatives, as were Tickellia and Abroscopus, but Abroscopus has not previously been considered to be closely related with Cettia and Tesia (Wolters, 1975–1982; Mayr and Cottrell, 1986; Sibley and Monroe, 1990; Inskipp et al., 1996; Dickinson, 2003). The study by Alström et al. (2006) of myoglobin intron II and mt-cytochrome b gene confirmed that Cettia was non-monophyletic and that there were near relationships among Cettia, Tesia, Urosphena, Abroscopus and Tickellia.
In this study, the Tesia and Abroscopus species grouped with four species from Cettia, forming a strongly supported clade (Figs. 1 and 2). The sequence divergence in cyt b between Abroscopus albogularis and Cettia species is from 0.121 (A. albogularis and C. diphone) to 0.151 (A. albogularis and C. flavolivaceus). However, the sequence divergence in cyt b between A. albogularis and the taxa of other genera in Sylviidae is from 0.131 to 0.192. Hence, the sequence divergences in cyt b between A. albogularis and Cettia species are smaller than those between A. albogularis and other generic species in Sylviidae. Similarly, the sequence divergences in cyt b between T. castaneocoronata and Cettia species are also smaller than those between T. castaneocoronata and other Sylviid genera. Although both trees depict Cettia as a monophyletic group, we await more study samples to clarify it as a monophyletic or non-monophyletic group.
Relations among other genera
Sibley and Monroe (1990) suggested that the Cisticolidae family included Prinia and Cisticola and that both Orthotomus and Locustella were placed into Acrocephalinae of Sylviidae. All the same, in some molecular studies, Prinia, Orthotomus and Cisticola have been found to be closely related, based on mitochondrial cyt b and 16S RNA (Cibois et al., 1999), mitochondrial ND2 and 12S RNA (Sefc et al., 2003) and nuclear RAG-1 and RAG-2 (Beresford et al., 2005). Nguembock et al. (2007) supported the placement of two Orthotomus species within the Cisticolidae. Alström et al. (2006) also supported this and suggested that Prinia, Orthotomus, Cisticola and other genera not studied here could be placed into Cisticolidae. Our results revealed a close relationship among Prinia, Orthotomus and Cisticola with good nodal support. Although Locustella species are clustered with a sister group comprising Prinia, Orthotomus and Cisticola in a terminal branch, the nodal support value derived by bootstrap of this clade is low in the maximum-likelihood analysis. Accordingly, more species from four genera Locustella, Prinia, Orthotomus and Cisticola are needed to resolve their evolutionary and phylogenetic relationships decisively.
Haffer (1991) suggested that Acrocephalus and Locustella are closely related, but that was disputed by Helbig and Seibold (1999). Leisler et al. (1997) and Helbig and Seibold (1999) proposed that Acrocephalus is non-monophyletic. In the present study, three members of Acrocephalus are clustered within the same clade with very high bootstrap and posterior probability. However, we have only a limited supply of samples and further extensive studies are therefore needed to review the taxonomic status and phylogeny of these two genera.
Acknowledgments
The support for this research was supplied by the National Science Funds for Distinguished Young Scientists (30925008) to F.M. Lei, the NSFC program (J0930004) to C.Y. Dai and the International Cooperation between the Chinese and Slovak Academy of Sciences to F.M. Lei and A. Krištín, as well as by the Program of the Ministry of Education of China (206148) to Z.M. Lian, the Program of the Education Department of the Shaanxi Provincial Government (2010JK909) and the Program of the Science and Technology Department of Yan'an, China (YAKY200701) to X. Lei. We thank Y. Huang for laboratory facilities and S.Q. Xu for data analysis.
Akaike H. 1973. Information theory as an extension of the maximum likelihood principle. In: Petrov BN, Csaki F (eds) Second International Symposium on Information Theory. Akademiai Kiadó, Budapest: 267–281.
Aliabadian M, Kaboli M, Nijman V, Vences M. 2009. Molecular identification of birds: performance of distance-based DNA barcoding in three genes to delimit parapatric species. PLoS ONE 4, e4119.
Alström P, Ericson PGP, Olsson U, Sundberg P. 2006. Phylogeny and classification of the avian superfamily Sylvioidea. Mol Phylogenet Evol, 38: 381–397.
Alström P, Olsson U, Rasmussen PC, Yao C-T, Ericson PGP, Sundberg P. 2007. Morphological, vocal and genetic divergence in the Cettia acanthizoides complex (Aves: Cettiidae). Zool J Linn Soc, 149: 437–452.
Alström P, Rasmussen PC, Olsson U, Sundberg P. 2008. Species delimitation based on multiple criteria: the Spotted Bush Warbler Bradypterus thoracicus complex (Aves: Megaluridae). Zool J Linn Soc, 154: 291–307.
Barker FK, Barrowclough GF, Groth JG. 2002. A phylogenetic hypothesis for passerine birds: taxonomic and biogeographic implications of an analysis of nuclear DNA sequence data. Proc Royal Soc Lond B, 269: 295–308.
Barker FK, Cibois A, Schikler P, Feinstein J, Cracraft J. 2004. Phylogeny and diversification of the largest avian radiation. Proc Natl Acad Sci USA, 101: 11040–11045.
Beresford P, Barker FK, Ryan PG, Crowe TM. 2005. African endemics span the tree of songbirds (Passeri): molecular systematics of several evolutionary 'enigmas'. Proc Royal Soc Lond B, 272: 849–858.
Cheng TH. 1994. A Complete Checklist of Species and Subspecies of the Chinese Birds. Science Press, Beijing. (in Chinese)
Cheng TH. 2000. A Complete Checklist of Species and Subspecies of the Chinese Birds (Revised Edition). Science Press, Beijing. (in Chinese)
Cibois A. 2003. Sylvia is a babbler: taxonomic implications for the families Sylviidae and Timaliidae. Bull Brit Ornithol Club, 123: 257–261.
Cibois A, Pasquet E, Schulenberg TS. 1999. Molecular systematics of the Malagasy babblers (Passeriformes: Timaliidae) and warblers (Passeriformes: Sylviidae), based on cytochrome b and 16S rRNA sequences. Mol Phylogenet Evol, 13: 581–595.
DeFilippis V. 1995. Evolution of the mitochondrial encoded cytochrome oxidase I gene versus the cytochrome b gene in ten species of Picidae. Master Thesis. Wayne State University, Detroit.
Dickinson EC, 2003. The Howard and Moore Complete Checklist of the Birds of the World. Christopher Helm, London.
Drovetski SV, Zink RM, Fadeev IV. 2004. Mitochondrial phylogeny of Locustella and related genera. J Avian Biol, 35: 105–110.
Ericson PGP, Johansson US. 2003. Phylogeny of Passerida (Aves: Passeriformes) based on nuclear and mitochondrial sequence data. Mol Phylogenet Evol, 29: 126–138.
Groth JG. 1998. Molecular phylogenetics of finches and sparrows: consequences of character state removal in cytochrome b sequences. Mol Phylogenet Evol, 10: 337–390.
Haffer J. 1991. Familie Sylviidae-Zweigsänger (Grassmücke und Verwandte). In: Glutz von Blotzheim UN, Bauer KM (eds) Handbuch der Vögel Mitteleuropas. Aula-Verlag, Wiesbaden. (in German)
Hebert PDN, Stoeckle MY, Zemlak TS, Francis CM. 2004. Identification of birds through DNA barcodes. PLoS Biol, 2: e312.
Helbig AJ, Seibold I. 1999. Molecular phylogeny of Palearctic–African Acrocephalus and Hippolais warblers (Aves: Sylviidae). Mol Phylogenet Evol, 11: 246–260.
Inskipp T, Lindsey N, Duckworth W. 1996. An Annotated Checklist of the Birds of the Oriental Region. Sandy, Bedfordshire.
Irwin DE, Alström P, Olsson U, Benowitz-Fredericks ZM. 2001. Cryptic species in the genus Phylloscopus (Old World leaf warblers). Ibis, 143: 233–247.
Jia CX, Sun Y-H, Bi ZL. 2003. The taxonomic status of Chinese Phylloscopus species. Acta Zootaxon Sin, 28: 202–209.
Johnson KP. 2001. Taxon sampling and the phylogenetic position of Passeriformes: Evidence from 916 avian cytochrome sequences. Syst Biol, 50: 128–136.
Klicka J, Fry AJ, Zink RM, Thompson CW. 2001, A cytochrome-b perspective on Passerina bunting relationships. Auk, 118: 610–623.
Kumar S, Tamura K, Nei M. 2004. MEGA3: Integrated software for molecular evolutionary genetics analysis and sequence alignment. Brief Bioinform, 5: 150–163.
La Touche JDD. 1925–1934. A Handbook of the Birds of Eastern China, Vol. 1, Passeriformes. Taylor and Francis, London.
Leache AD, Reeder TW. 2002. Molecular systematics of the eastern fence lizard (Sceloporus undulatus): a comparison of parsimony, likelihood, and Bayesian approaches. Syst Biol, 51: 44–68.
Leisler B, Heidrich P, Schulze-Hagen K, Wink M. 1997. Taxonomy and phylogeny of reed warblers (genus Acrocephalus) based on mtDNA sequences and morphology. J Ornithol, 138: 469–496.
MacKinnon JR, Phillipps K. 2000. A Field Guide to the Birds of China. Oxford University Press, Oxford.
Martens J, Sun Y-H, Päckert M. 2008. Intraspecific differentiation of Sino-Himalayan bush-dwelling Phylloscopus warblers, with description of two new taxa (P. fuscatus, P. fuligiventer, P. affinis, P. armandii, P. subaffinis). Vert Zool, 58: 233–265.
Mayr E, Cottrell GW. 1986. Checklist to the Birds of the World, Vol. XI. Museum of Comparative Zoology, Cambridge.
Monroe Jr BL, Sibley CG. 1993. A World Checklist of Birds. Yale University Press, New Haven & London.
Nguembock B, Fjeldså J, Tillier A, Pasquet E. 2007. A phylogeny for the Cisticolidae (Aves: Passeriformes) based on nuclear and mitochondrial DNA sequence data, and a re-interpretation of an unique nest-building specialization. Mol Phylogenet Evol, 42: 272–286.
Olsson U, Alström P, Sundberg P. 2004. Non-monophyly of the avian genus Seicercus (Aves: Sylviidae) revealed by mitochondrial DNA. Zool Scr, 33: 501–510.
Olsson U, Alström P, Ericson PGP, Sundberg P. 2005. Non-monophyletic taxa and cryptic species — evidence from a molecular phylogeny of leaf-warblers (Phylloscopus, Aves). Mol Phylogenet Evol, 36: 261–276.
Päckert M, Blume C, Sun Y-H, Wei L, Martens J. 2009. Acoustic differentiation reflects mitochondrial lineages in Blyth's leaf warbler and white-tailed leaf warbler complexes (Aves: Phylloscopus reguloides, Phylloscopus davisoni). Biol J Linn Soc, 96: 584–600.
Pasquet E, Cibois A, Baillon F, Erard C. 2002. What are African monarchs (Aves, Passeriformes)? A phylogenetic analysis of mitochondrial genes. C.R. Biologies, 325: 107–118.
Posada D, Crandall KA. 1998. MODELTEST: testing the model of DNA substitution. Bioinformatics, 14: 817–818.
Saetre GP, Borge T, Lindell J, Moum T, Primmer CR, Sheldon BC, Haavie J, Johnsen A, Ellegren H. 2001. Speciation, introgressive hybridization and nonlinear rate of molecular evolution in flycatchers. Mol Ecol, 10: 737–749.
de Schauensis RM. 1984. The Birds of China. Smithsonian Institution Press, Washington D.C.
Sefc KM, Payne RB, Sorenson MD. 2003. Phylogenetic relationships of African sunbird-like warblers: Moho (Hypergus atriceps), Green Hylia (Hylia prasina) and Tit-hylia (Pholidornis rushiae). Ostrich, 74: 8–17.
Sheldon FH, Whittingham LA, Moyle RG, Slikas B, Winkler DW. 2005. Phylogeny of swallows (Aves: Hirundinidae) estimated from nuclear and mitochondrial DNA sequences. Phylogenet Evol, 35: 254–270.
Sibley CG, Ahlquist JE. 1990. Phylogeny and Classification of Birds. Yale University Press, New Haven.
Sibley CG, Monroe Jr BL. 1990 Distribution and Taxonomy of Birds of the World. Yale University Press, New Haven.
Sorenson MD, Ast JC, Dimcheff DE, Yuri T, Mindell DP. 1999. Primers for a PCR-based approach to mitochondrial genome sequencing in birds and other vertebrates. Mol Phylogenet Evol, 12: 105–114.
Swofford DL. 2002. PAUP*: Phylogenetic analysis using parsimony (* and other methods), Version 4.0b10. Sinauer Associates, Sunderland, Massachusetts.
Thomassen HA, Wiersema AT, de Bakker MA, de Knijff P, Hetebrij E, Povel GD. 2003. A new phylogeny of swiftlets (Aves: Apodidae) based on cytochrome-b DNA. Mol Phylogenet Evol, 29: 86–93.
Thompson JD, Gibson TJ, Plewniak F, Jeanmougin F, Higgins DG. 1997. The Clustal X windows interface: flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res, 24: 4876–4882.
Ticehurst CB. 1938. A Systematic Review of the Genus Phylloscopus. Trustees of the British Museum, London.
Vaurie C. 1965. The Birds of the Palearctic Fauna. Order Passeriformes. H. F. and G. Witherby Ltd., London.
Watson GE, Traylor MA, Mayr E. 1986. Family Sylviidae, Old World warblers. In: Mayr E, Cottrell GW (eds) Checklist to the Birds of the World, Vol XI. Museum of Comparative Zoology, Cambridge.
Webb DM, Moore WS. 2005. A phylogenetic analysis of woodpeckers and their allies using 12S, Cyt b and COI nucleotide sequences (class Aves; order Piciformes). Mol Phylogenet Evol, 36: 233–248.
Weibel AC, Moore WS. 2002. Molecular phylogeny of a cosmopolitan group of woodpeckers (Genus Picoides) based on COI and Cyt b mitochondrial gene sequences. Mol Phylogenet Evol, 22: 65–75.
Wolters HE. 1975–1982. Die Vogelarten der Erde. Paul Parey, Hamburg & Berlin.
Zheng GM. 2005. A Checklist on the Classification and Distribution of the Birds of China. Science Press, Beijing. (in Chinese)

 DownLoad:
DownLoad:





 Email Alerts
Email Alerts RSS Feeds
RSS Feeds