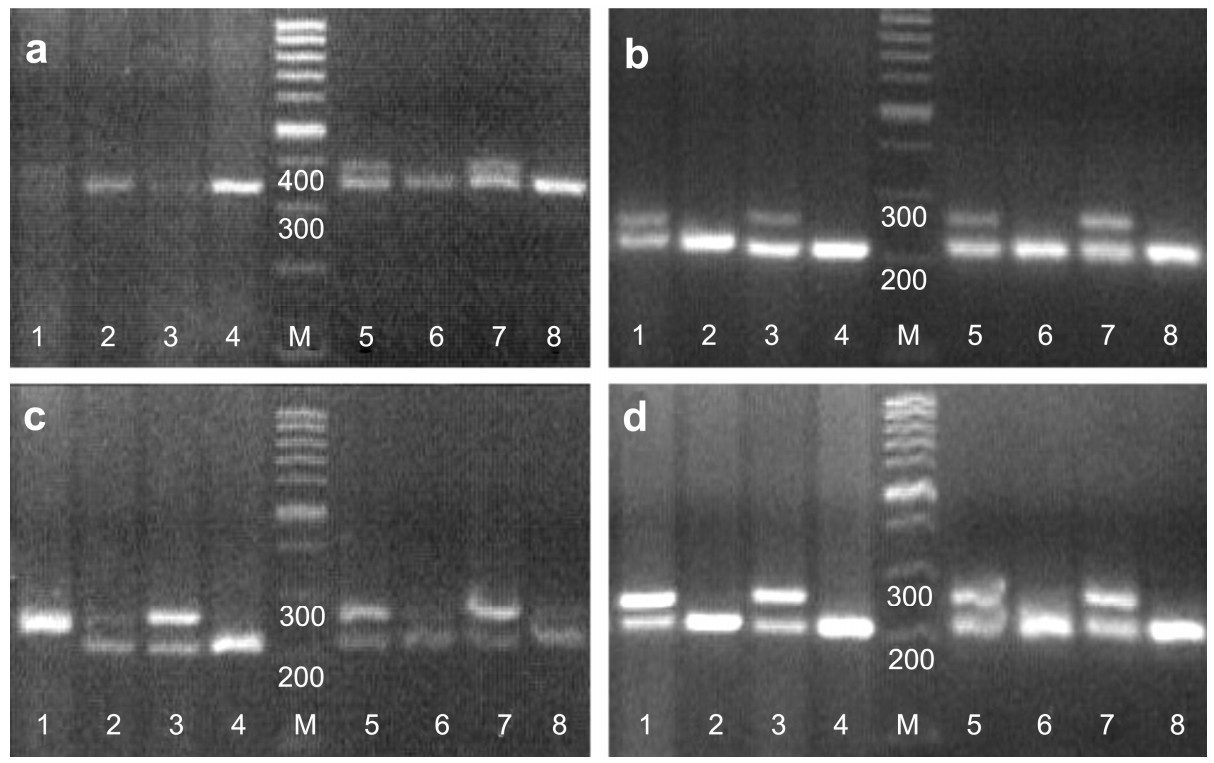
The primer pair sex1/sex2, which can be widely applied for sex identification in Galliform species, was used to determine the sex of 17 Passeriform species. As CHD-W fragments tended to be preferentially amplified, which may cause unnecessary misidentification in bird species with little difference between CHD-Z and CHD-W, we modified sex1 and sex2, obtaining sex1' and sex-mix respectively. Primer sets were then recombined to conduct sex identification. After testing several Passeriforme birds of known sex, we found that the primer pair sex1'/sex2 was better at limiting the preferential amplification of CHD-W fragments. As they are being frequently used in sex allocation study of Aegithalos concinnus and song learning research of Lonchura striata, we can expect more applications of this primer pair to further studies in Passeriformes.
Global biodiversity is facing increasing threats due to climate change and anthropogenic impacts (Myers et al., 2000; Grenyer et al., 2006). Identification of important species, under risk of extinction, would be of great help to biologists in a more efficient allocation of limited funds to improve and conserve biological diversity by focusing on these unique species (Grenyer et al., 2006). A common practice to identify conservation values of species is to evaluate the rarity of the population of a species, its distribution ranges (He, 2012) and changes in habitat conditions as is practised in the IUCN Red List (The World Conservation Union, 2010). In recent years, the importance of evolutionary history has become recognized (Cadotte and Davies, 2010; Martyn et al., 2012) and many phylogenetic diversity (PD) indices for quantifying evolutionary heritage of species have been proposed in a number of studies (Faith, 1992, 2002; Faith et al., 2004; Tucker et al., 2012).
The avifauna of China is composed of around 1314 species of which 52 are said to be endemic (only found within the political boundaries of China, i.e., the mainland and Taiwan) (Lei and Lu, 2006). Conservation priorities of these endemic and other bird species in China have been widely established in previous studies (Lei et al., 2002a, 2007; Chen, 2007, 2008; Chang et al., 2013). However, the importance of conservation of endemic birds of China from a phylogenetic perspective has never been evaluated up till now. As such, the present report presents a way to fill such a knowledge gap by proposing conservation priority of endemic birds of China using a series of phylogenetic diversity metrics.
Table
1.
Conservation priorities of species based on alternative phylogenetic diversity metrics. IUCN categories: EN (endangered), VU (vulnerable), NT (near threatened), LC (least concerned), DD (data deficient). "-" denotes the IUCN information is not available. Codes for phylogenetic diversity metrics: ES (equal split), FP (fair proportions), ED (evolutionary distinctiveness), TD (taxonomic distinctiveness), PL (pendant edge's length) and Node-based I and W indices.
The phylogenetic relationships between the 52 endemic birds of China's mainland are extracted from the BirdTree.org database (http://www.birdtree.org), which is derived from a full phylogeny of the global bird species in a previous study (Jetz et al., 2012). However one species, Ficedula beijingnic, was omitted from the tree files of the database. It was therefore decided to exclude this species for further analyses, while the remaining 51 species were used for analysis since these are all included in the retrieved phylogenetic trees. From the 3000 trees tested for possible phylogenetic affinities, the 51 endemic birds were retrieved and the resultant consensus tree, with average branch lengths, was obtained using the DendroPy python library (Sukumaran and Holder, 2010). Molecular dating of the tree was carried out using a penalized likelihood method (Sanderson, 2002). The result, a dated tree (Fig. 1), is used for all subsequent analyses.
Figure
1.
Consensus phylogenetic tree for 51 endemic birds of mainland China
As a comparison, the category of the IUCN Red list for each species was collected from BirdLife International (http://www.birdlife.org/) and included in Table 1. The abbreviations for each category are as follows: EN (endangered), VU (vulnerable), NT (near threatened), LC (least concerned) and DD (data deficient). When comparing the rankings of IUCN and PD indices, the IUCN categories are transformed into discrete integers, i.e., EN (1), VU (2), NT (3), LC (4) and DD (5). The Wilcox signed rank test and Pearson's correlation test were implemented to compare both rankings. A significant Wilcox signed rank or a non-significant Pearson's correlation coefficient implies that both rankings are fundamentally different.
Results
As shown in Table 1, the top five endemic birds based on the combined rankings of seven PD indices in order of priority, are Arborophila ardens, A. gingica, A. rufipectus, Lophophorus lhuysii and Alectoris magna. Their corresponding IUCN classes are VU, NT, EN, VU and LC, respectively. As seen, the PD-based ranking accurately identified the conservation importance of Arborophilarufipectus, an endangered species that should have a high priority for conservation.
Arborophila rufipectus has a very limited range of distribution in the southern part of Sichuan Province. The size of its adult population is estimated to be around 1000 (BirdLife International, http://www.birdlife.org/). Due to continuous hunting and habitat loss, it was classified as Critically Endangered in previous IUCN reports(1994, 1998). It is now still listed in the category of Endangered species. Based on a phylogenetic perspective this species, along with two other Arborophila species, has a very unique and long evolutionary history, when compared to other endemic birds (Fig. 1).
The PD-ranking for Arborophila ardens and Lophophorus lhuysii are also fairly accurate (top-1 and 4), both of which are listed as VU in the IUCN Red List (The World Conservation Union, 2010). This implies that both should deserve high conservation priority from an ecological perspective (based on IUCN ranking) as well as from an evolutionary (based on PD ranking) perspective.
In contrast, there are some inconsistencies between PD and IUCN rankings. For example, Arborophilagingica and Alectoris magna are ranked 2nd and 5th in the PD ranking for their unique evolutionary histories. However, in the IUCN ranking, these species are merely grouped into the categories of NT and LC.
Overall, the difference of rankings based on the IUCN Red List and the combined PD ranking is statistically significant (Wilcox signed rank test: V = 159, p < 0.001; Pearson's correlation coefficient: r = 0.217, p = 0.129). These tests show a significant difference between IUCN and PD rankings for endemic birds of mainland China.
Discussion
Conservation importance of endemic or rare species has been widely recognized (Linder, 1995; Lamoreux et al., 2006; Gaston, 2012). In the present study, I quantified the conservation importance of avian species, endemic to China, by utilizing a variety of phylogenetic diversity metrics. The results show a statistically significant difference between the priority rankings based on PD metrics and that derived from the IUCN Red list. Therefore, a PD-based conservation emphasis on endemic birds of China might offer some new views when establishing relevant conservation strategies by considering evolutionary heritage and genetic resources of these species.
The present study carried out analyses on 51 endemic birds. However, it might be a bit ambiguous given the number of endemic birds in China (Zhang, 2004), when one considers migration during the breeding season (Lei et al., 2002b). In a previous study, the number of endemic birds in China was believed to be around 100 (Lei et al., 2002a), while in a more recent publication, this number is said to be 105 (Lei and Lu, 2006). If the definition of endemic birds were extended to include, for example, Taiwan and Hong Kong, the number of endemic birds should be at least 70 (Zhang, 2004; Lei et al., 2002b). When considering only mainland China, the number of endemic birds should be at least 50, based on the information from the World Bird Database (http://avibase.bsc-eoc.org/). Therefore, the present study should be relatively accurate in suggesting conservation priorities for endemic birds in mainland China based on the phylogenetic diversity framework. Any re-analyses with the addition of a few more possible endemic birds should not greatly affect the quantitative results presented in the present study.
In this study, I have only used pure phylogeny-based diversity indices without considering other weighted phylogenetic diversity indices. Weighted phylogenetic diversity metrics such as the abundance-weighted PD index (Cadotte et al., 2010), the biogeography-weighted PD index (Tucker et al., 2012), the endemism-weighted PD index (Rosauer et al., 2009) and possibly other indices might provide more insights into conservation priorities because these can explicitly incorporate the role of some biological factors when studying the evolutionary history of species. For future implications, integration of other weighted phylogenetic diversity indices into the systematic conservation planning of birds would offer new insights and thus should be considered in the next levels of research.
Acknowledgements
This work was supported by the University of British Columbia and now supported by China Scholarship Council.
Bensch S, Westerdahl H, Hansson B, Hasselquist D. 1999. Do females adjust the sex of their offspring in relation to the breeding sex ratio? J Evol Biol, 12: 1104–1109
Cockburn A, Double MC. 2008. Cooperatively breeding superb fairy-wrens show no facultative manipulation of offspring sex ratio despite plausible benefits. Behav Ecol Sociobiol, 62: 681–688
Dreiss A, Richard M, Moyen F, White J, Moller AP, Danchin E. 2006. Sex ratio and male sexual characters in a population of blue tits, Parus caeruleus. Behav Ecol, 17: 13–19
Dubiec A, Zagalska-Neubauer M. 2006. Molecular techniques for sex identification in birds. Biol Lett, 43: 3–12
Ellegren H. 1996. First gene on the avian W chromosome (CHD) provides a tag for universal sexing of non-ratite birds. Proc R Soc Lond B, 263: 1635–1641
Ellegren H, Sheldon BC. 1997. New tools for sex identification and the study of sex allocation in birds. Trends Ecol Evol, 12: 255–259
Ellegren H. 2002. Dosage compensation: do birds do it as well? Trends Genet, 18: 25–28
Fiala KL. 1980. On estimating the primary sex ratio from incomplete data. Am Nat, 115: 442–444
Frank SA. 1990. Sex allocation theory for birds and mammals. Ann Rev Ecol Syst, 21: 13–55
Fridolfsson AK, Ellegren H. 1999. A simple and universal method for molecular sexing of non-ratite birds. J Avian Biol, 30: 116–121
Griffiths R, Double MC, Orr K, Dawson RJG. 1998. A DNA test to sex most birds. Mol Ecol, 7: 1071–1075
Hatchwell BJ, Russell AF. 1996. Provisioning rules in cooperatively breeding long-tailed tits Aegithalos caudatus: An experimental study. Proc R Soc Lond B Biol Sci, 263: 83–88
Kahn NW, John JS, Quinn TW. 1998. Chromosome-specific intron size differences in the avian CHD gene provide an efficient method for sex identification in birds. Auk, 115: 1074–1078
Lessells K, Mateman C. 1996. Molecular sexing of birds. Nature, 383: 761–762
McGowan A, Hatchwell BJ, Woodburn RJW. 2003. The effect of helping behaviour on the survival of juvenile and adult long-tailed tits Aegithalos caudatus. J Anim Ecol, 72: 491–499
McQueen HA, McBride D, Miele G, Bird AP, Clinton M. 2001. Dosage compensation in birds. Curr Biol, 11: 253–257
Nakagawa S. 2004. Is avian sex determination unique?: clues from a warbler and from chickens. Trends Genet, 20: 479–480
Quinn TW, Cooke F, White BN. 1990. Molecular sexing of geese using a cloned Z chromosomal sequence with homology to the W chromosome. Auk, 107: 199–202
Saino N, de Ayala RM, Martinelli R, Boncoraglio G. 2008. Male-biased brood sex ratio depresses average phenotypic quality of barn swallow nestlings under experimentally harsh conditions. Oecologia, 156: 441–453
Wang N, Zhang ZW. 2009. The novel primers for sex identification in the brown eared-pheasant and their application to other species. Mol Ecol Resour, 9: 186–188
West SA, Sheldon BC. 2002. Constraints in the evolution of sex ratio adjustment. Science, 295: 1685–1688
Whittingham LA, Dunn PO. 2000. Offspring sex ratios in tree swallows: females in better condition produce more sons. Mol Ecol, 9: 1123–1129
Woxvold IA, Magrath MJL. 2008. Sex-biases in the hatching sequence of cooperatively breeding apostlebirds Struthidea cinerea. Evol Ecol, 22: 139–151
Table
1.
Conservation priorities of species based on alternative phylogenetic diversity metrics. IUCN categories: EN (endangered), VU (vulnerable), NT (near threatened), LC (least concerned), DD (data deficient). "-" denotes the IUCN information is not available. Codes for phylogenetic diversity metrics: ES (equal split), FP (fair proportions), ED (evolutionary distinctiveness), TD (taxonomic distinctiveness), PL (pendant edge's length) and Node-based I and W indices.

 DownLoad:
DownLoad:






 Email Alerts
Email Alerts RSS Feeds
RSS Feeds