Peter SAMAŠ, Lenka POLAČIKOVÁ, Mark E. HAUBER, Phillip CASSEY, Tomáš GRIM. 2012: Egg rejection behavior and clutch characteristics of the European Greenfinch introduced to New Zealand. Avian Research, 3(4): 330-338. DOI: 10.5122/cbirds.2012.0037
Citation:
Peter SAMAŠ, Lenka POLAČIKOVÁ, Mark E. HAUBER, Phillip CASSEY, Tomáš GRIM. 2012: Egg rejection behavior and clutch characteristics of the European Greenfinch introduced to New Zealand. Avian Research, 3(4): 330-338. DOI: 10.5122/cbirds.2012.0037
Peter SAMAŠ, Lenka POLAČIKOVÁ, Mark E. HAUBER, Phillip CASSEY, Tomáš GRIM. 2012: Egg rejection behavior and clutch characteristics of the European Greenfinch introduced to New Zealand. Avian Research, 3(4): 330-338. DOI: 10.5122/cbirds.2012.0037
Citation:
Peter SAMAŠ, Lenka POLAČIKOVÁ, Mark E. HAUBER, Phillip CASSEY, Tomáš GRIM. 2012: Egg rejection behavior and clutch characteristics of the European Greenfinch introduced to New Zealand. Avian Research, 3(4): 330-338. DOI: 10.5122/cbirds.2012.0037
Department of Zoology and Laboratory of Ornithology, Palacký University, 17. listopadu 50, CZ-771 46 Olomouc, Czech Republic
2.
Department of Pathology and Parasitology, Faculty of Veterinary Medicine, University of Veterinary and Pharmaceutical Sciences, Palackého 1-3, CZ-612 42 Brno, Czech Republic
3.
Department of Psychology, Hunter College and the Graduate Center of the City University of New York, 695 Park Avenue, New York, NY 10065, USA
4.
School of Earth and Environmental Sciences, University of Adelaide, SA 5005, Australia
Animal populations, with a known history of introduction events, provide opportunities to study the dynamics of how rapid shifts in ecological context affect behavioral (e.g., responses to brood parasitism) and life-history (e.g., clutch and egg parameters) traits. We studied the European Greenfinch (Carduelis chloris) introduced to New Zealand, regarding foreign-egg rejection behaviors and also compared their clutch characteristics with data from the source populations in the United Kingdom. Although previously this species had been considered an unsuitable host for the Common Cuckoo (Cuculus canorus), and not impacted by selection pressure associated with brood parasitism, we found that Greenfinches in our study population were able to eject experimental eggs at low frequencies. In contrast, nest desertion rates were similar in experimentally parasitized and control unmanipulated nests, implying that nest desertion is not an antiparasite adaptation in this species. Contrary to previous studies, we did not find significant differences in clutch and egg sizes between introduced and source populations. This study emphasizes (1) the importance of using control treatments in studies of host responses to experimental parasitism, (2) including apparently unsuitable hosts of brood parasites, and (3) meta-replicating prior studies to further the process of gaining and validating scientific knowledge.
Avian brood parasitism provides a powerful study system with challenging questions regarding the direction and speed of coevolutionary arms races, including host selection, antagonistic interspecies interactions, and perceptual mimicry (Davies, 2011; Kilner and Langmore, 2011). Obligate brood parasitic birds, including the Common Cuckoo (Cuculus canorus; hereafter: Cuckoo), and their hosts provide ideal models to study strong reciprocal selection pressures, because parasitism imposes moderate-to-severe costs on host parents that provision unrelated young in their nests and hosts evolve antiparasitic traits (Davies, 2011). Such coevolutionary arms races have resulted in a diverse set of quantifiable differences in the behaviors of both typical and potential host populations that are either in contact, or in isolation, with a brood parasite (Davies and Brooke, 1989; Lahti, 2005; Hale and Briskie, 2007; Stokke et al., 2007; Grim et al., 2011).
Research effort devoted to brood parasitism in birds has increased dramatically in recent decades (Grim, 2007). However, most studies have focused only on suitable Cuckoo hosts (Davies, 2011), whereas antiparasitic behaviors of rare (Moksnes and Røskaft, 1992; Honza et al., 2004; Procházka and Honza, 2004) or unsuitable (Davies and Brooke, 1989; Moksnes et al., 1991; Grim et al., 2011) hosts have been tested only occasionally. Studying apparently unsuitable Cuckoo hosts provides important insights for our understanding of hostparasite coevolution. For example, variation in the behavioral responses of unsuitable hosts to experimental parasitism can simulate conditions at the start of brood parasitism in a novel (i.e., so far unused) host, and also give information on the phylogenetic distribution of behavioral traits that can serve as pre-adaptations for defenses against brood parasites (Moskát et al., 2003a).
Species which have some allopatric populations, and other sympatric populations, with brood parasites, provide valuable opportunities to study potential specieslevel flexibility of antiparasitic strategies (Davies and Brooke, 1989; Stokke et al., 2007; Grim et al., 2011). Importantly, almost all studies on the effects of sympatry vs. allopatry with a brood parasite on host responses were conducted in regions for which the length of sympatry or allopatry was unknown, i.e., reliable historical information on how long parasites were absent or cohabiting with hosts is missing in most cases (Davies and Brooke, 1989; Røskaft et al., 2002; Grim et al., 2011). Rare exceptions include the work of Lahti (2005) who knew accurately the length of allopatry in populations of Village Weaverbirds (Ploceus cucullatus) introduced from Africa to the islands of Mauritius and Hispaniola, and Hale and Briskie (2007) who knew the exact length of allopatry in populations of several European avian species introduced to New Zealand (see also Samaš et al., 2011).
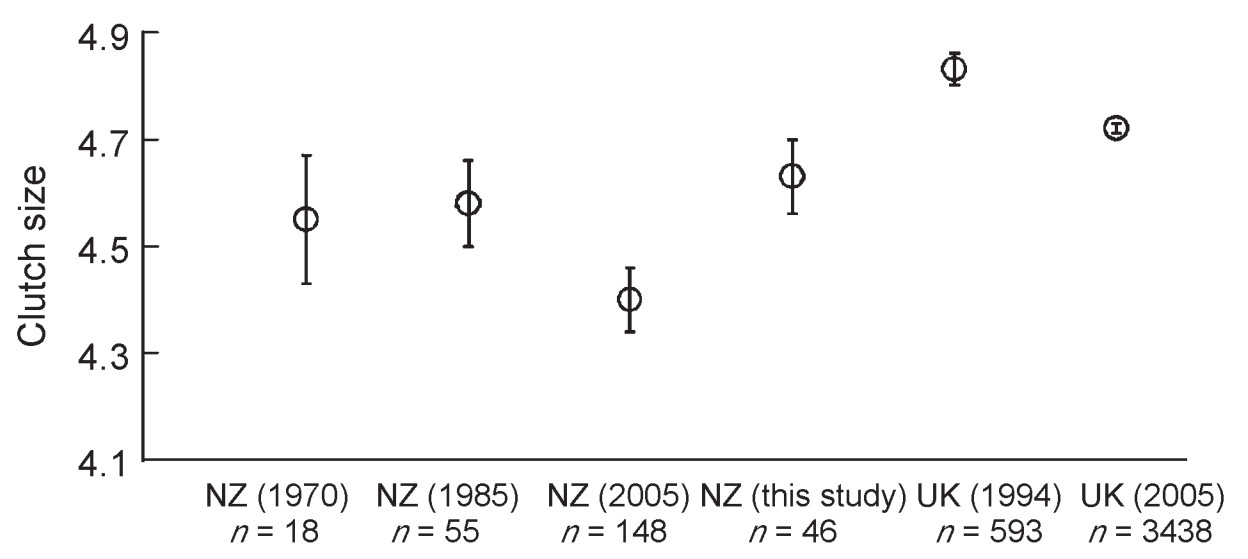
Here, we take advantage of the known length of complete allopatry with Cuckoos in the European Greenfinch (Carduelis chloris; hereafter: Greenfinch); this species was introduced from the United Kingdom (hereafter: UK) to New Zealand (hereafter: NZ) between 1863–1868 (Thomson, 1922). Although less than 100 individuals were introduced, today the Greenfinch is common throughout NZ and there is no genetic evidence for severe inbreeding or genetic drift in this species (Merilä et al., 1996; see also Briskie and MacKintosh, 2004). This is important as bottlenecks could lead to genetic changes and may significantly influence clutch characteristics, behavioral and sensory traits and other aspects of introduced populations (Briskie and MacKintosh, 2004; Congdon and Briskie, 2010). In contrast, several studies have detected divergent changes in the breeding characteristics of the Greenfinch between the source population in the UK and the introduced population in NZ, including lower clutch size (Niethammer, 1970; MacMillan, 1985; Evans et al., 2005) and smaller egg size (Cassey et al., 2005) in NZ. These changes support general observations that passerines in the Northern Hemisphere have higher clutch sizes, fewer breeding attempts per season and/or a shorter breeding season in contrast to the Southern Hemisphere regions (Martin, 1996; Samaš et al., 2013).
Nevertheless, methodologically independent replication of whole studies (so called "meta-replication") is critically important for the validity and generality of scientific information (Johnson, 2002; Kelly, 2006). Therefore, additionally to testing Greenfinch responses to experimental brood parasitism in allopatry with the Cuckoo, we also studied critical parameters of Greenfinch breeding biology in NZ (clutch and egg sizes). Traits of reproductive investment, including clutch size and egg size, are important components of avian parasite-host arms races because clutch and egg sizes may be related to individual host selection by female parasites and egg rejection decisions by hosts, and clutch size may evolve divergently between host populations sympatric vs. allopatric with parasites (e.g., Soler et al., 2001; Hauber, 2003; Servedio and Hauber, 2006; White et al., 2007). Increase of clutch size might also act as a defensive trait against brood parasites. For example, Svensson and Råberg (2010) and Soler et al. (2011) suggested that individual hosts better tolerate interspecific parasitism by laying larger clutches.
The Greenfinch is considered an unsuitable host for the Cuckoo because it feeds its nestlings mostly with plant seeds (Davies and Brooke, 1989; Moksnes et al., 1991). However, expectations on host suitability based on nestling diet composition have been shown empirically to be misleading in certain cases (Martín-Gálvez et al., 2005; Grim, 2006). Indeed, there are several documented reports of Cuckoo chicks being successfully raised by the Greenfinch (Seel and Davis, 1981). Clearly, more studies are needed to understand GreenfinchCuckoo interactions in the context of brood parasitism, for example, to test whether flexible foraging and diet selection by Greenfinches, the flexibility of young Cuck oos digestion, or the interaction of both these factors can explain variation in the Cuckoo chick's potential survival in Greenfinch nests (see also Grim et al., 2011).
No study has specifically tested the behavior of Greenfinch regarding egg rejection behavior in NZ (cf. Boulton and Cassey, 2006; Hale and Briskie, 2007). In addition, only two European studies have reported responses of the Greenfinch to experimental brood parasitism. Davies and Brooke (1989) used three types of hard and solid artificial egg models that were painted to resemble eggs of Cuckoos parasitizing Common Redstarts (Phoenicurus phoenicurus), Pied Wagtails (Motacilla alba) and Meadow Pipits (Anthus pratensis). None of these models introduced during the laying or incubation stages (n = 12) were rejected by Greenfinches (Table 3 in Davies and Brooke, 1989). However, when a Meadow Pipit-like model egg was placed in completed Greenfinch nests before laying began, 2 out of 7 Greenfinches showed rejection (1 deserted, 1 built over; p. 216 in Davies and Brooke, 1989). Thus, the Greenfinch in the UK seems to be capable of responding to foreign eggs in the nest at least in some circumstances.
Elsewhere, Moksnes et al. (1991) reported that 41% Greenfinches (n = 17) rejected non-mimetic model eggs when introduced at both laying and incubation stages. Rejections (n = 7; Moksnes et al., 1991) were mostly by desertion (n = 5) which may or may not have been a specific response to the appearance of a foreign egg (Kosciuch et al., 2006; see also Results of the present study). Other responses were described as "unselective ejection" (n = 2) which means that "one, several, or all of the host's eggs were destroyed or removed. In the majority of such cases, the artificial egg was left in the nest" (p. 350 in Moksnes et al., 1991). Such "unselective ejection" would be difficult to distinguish from the impacts of nest predators, especially if all natural, host eggs disappear or are destroyed (see Weidinger, 2010). Furthermore, as both of these previous studies were conducted without a control non-manipulated nestor clutch-treatment, it remains unclear whether tested Greenfinches responded specifically to brood parasite eggs or non-specifically to a general clutch disturbance (see Kosciuch et al., 2006).
Here we experimentally tested egg rejection behavior in an introduced Greenfinch population in NZ. In addition, we compared egg and clutch sizes of this introduced population and their European source population. We predicted that the Greenfinch in NZ would not discriminate against alien eggs (Davies and Brooke, 1989), and show smaller clutch sizes and/or smaller egg sizes in NZ in the southern hemisphere than in northern UK source populations (Cassey et al., 2005).
Methods
All breeding and experimental data were collected during October and November 2007–2008 in the city of Hamilton in NZ and its rural vicinity (37°46'S, 175°16' E). We searched for Greenfinch nests in shrub vegetation and in plantations of blueberry (Vaccinium sp.) orchards. Individuals were not ringed for identification in this study. We used each nest as an independent data point in the statistical analyses.
We checked nests (n = 41) every 1–5 days. We marked all eggs (n = 158) on the blunt pole for individual recognition. After clutch completion we measured egg length and width by digital calliper to the nearest 0.01 mm.
For Greenfinches, the grasp or puncture ejection of hard artificial model may represent a problem due to beak size and geometry (Martín-Vivaldi et al., 2002). Therefore, we did not use hard plastic models but painted one randomly selected own host egg with a nontoxic SharpieTM felt tip black pen, following methods of Hauber et al. (2006), so that the experimental design allowed the "foreign" eggs to be puncture ejected. We painted the eggs black given that the natural colour of Greenfinch eggs is greyish-white to bluish-white, or beige, sparsely spotted dark reddish, concentrated at the blunt end (Cramp, 1994; see Fig. 1b in Boulton and Cassey, 2006). Thus, our experimental manipulation of the egg phenotype caused a large change in the host egg's appearance (chroma, brightness, and maculation) to provide a strong test of host egg rejection abilities compared to the potential use of mimetic experimental eggs (Grim, 2005). After painting the eggs we checked that there were no cracks in the eggshells that also might cause egg ejection or nest desertion and did not find any such cracks. Importantly, our manipulation did not alter the overall clutch size of the potential host (similarly to natural Cuckoo behavior of removing a host egg and replacing it with a parasitic egg; Davies, 2011). Egg painting had no effect on painted eggs' hatchability or chick survival at acceptor Greenfinch nests in our study population. Specifically, all painted eggs successfully hatched, chicks grew normally and we did not find any single missing chicks in manipulated nests (if chicks disappeared, it was always a whole brood, i.e., consequence of nest predation, see Weidinger, 2010).
We considered host reaction as an "acceptance" if the manipulated egg was still found in an active nest on the 6th day, or later (Davies and Brooke, 1989; Moksnes et al., 1991). If a manipulated egg disappeared within the 6-day period, but hosts continued incubating the clutch, we considered host reaction as "ejection". The presence of traces of yolk at remaining host eggs was considered as "puncture ejection" of ejected egg (following Moksnes et al., 1994; Antonov et al., 2006). Non-predated nests with eggs found to be unincubated (cold) during several consecutive nest checks were scored as "deserted" (Kosciuch et al., 2006). Finally, depredated nests were excluded from analyses.
Nest desertion may result from many causes (e.g., inclement weather, human disturbance, predator visit; Weidinger, 2010) and, thus, not always be a specific response to parasitism (Kosciuch et al., 2006). Therefore, it is also necessary to study host behavior at non-parasitized, control nests. Overall, we randomly assigned 20 nests as controls and another 20 nests as experimental treatments. In one additional nest we knew clutch size after clutch completion but the nest was predated during 4th day of experiment. We included this nest in analyses of clutch and egg sizes but had to exclude it from egg experimental treatment analyses, hence sample sizes differ between analyses. During the laying stage, we found and monitored three experimental and 10 control nests without predation, resulting in lower sample sizes for some specific analyses (see also Discussion).
Except for the egg phenotype manipulation, all nests were treated in the same way. We painted the experimental host own egg once the nest found active with two or more eggs. Painting lasted ~1 minute. Eggs were measured after clutch completion and were returned to nests immediately after measurement. During each nest visit we used an inspection mirror to check nest content and pulled out a random egg from a nest to check the egg temperature by touch (warm, i.e., incubated, or cold, i.e., deserted).
We obtained UK clutch size data for the Greenfinch from the national record card schemes organized by the BTO (Robinson, 2005) and egg sizes from Cramp et al. (1994), particularly for the UK endemic subspecies (Carduelis chloris harrisoni). To avoid pseudoreplication in egg length and width analyses, we used clutch averages as units of analysis (Cassey et al., 2005). We used Welsch's t-test for unequal variances in statistical testing of mean differences of clutch and egg sizes between NZ and UK (see Ruxton, 2006).
Results are shown as mean ±S.D. Statistical analyses were performed in JMP 8.0.1 (SAS Institute, 2009).
Results
Nest desertion
Data pooled across both breeding stages (laying + incubation) showed the same desertion rates of 20% (n = 4 out of 20) in both control and experimental nests. For control nests, there was no statistically significant difference in desertion rates between laying (3 out of 10) and incubation stages (1 out of 10; Fisher's exact test, twotailed, p = 0.31). The analogous comparison of experimental nests between laying (2 out of 3) and incubation stages (2 out of 17) also showed no statistical difference in desertion rates (Fisher's exact test, two-tailed, p = 0.09).
For the laying stage, desertion rates were similar in control (3 out of 10) and experimental nests (2 out of 3; Fisher's exact test, two-tailed, p = 0.51). Similarly, desertion rates during incubation were statistically similar between control (1 out of 10) and experimental nests (2 out of 17; Fisher's exact test, two-tailed, p = 1.00). Within experimental nests, clutch sizes at deserted nests were statistically similar as those at nests where Greenfinches either accepted or ejected the experimental egg (Table 1). Within control nests, clutch sizes did not differ between deserted and non-deserted nests (Table 1).
Table
1.
Clutch size (mean ±S.E.) of New Zealand introduced Greenfinches according to their behaviour at experimental and control nests. Differences within the experimental nest category were tested with Kruskal-Wallis test, all other tests were Welsch's t-tests for unequal variances (as recommended by Ruxton, 2006). Sample sizes slightly differ from those in Table 2 because of missing data for one experimental and one control nest.
At 2 out of 20 nests owners ejected painted eggs and in 1 case ejected own unpainted egg while the painted egg remained in nest (i.e., a case of rejection error, see Discussion; Table 2). Thus, hosts at 3 out of 20 nests showed egg ejection behavior. All 3 ejections were presumably by puncture ejection as evidenced by traces of yolk on the remaining eggs. Ejections occurred 1 to 3 days after the start of experiment (i.e., manipulation of the egg phenotype), respectively. Ejection rates at experimental (3 out of 20 nests) and control nests (0 out of 20) were not statistically different (Fisher's exact test, two-tailed, p = 0.23). Ejection rates at experimental nests did not differ between laying (1 out of 3 nests) and incubation stages (2 out of 17 nests; Fisher's exact test, two-tailed, p = 0.40).
Table
2.
Overview of New Zealand introduced Greenfinch responses to experimental parasitism (1 host own egg painted black) and control nest treatments. In all, we recorded 3 ejections, 2 cases of "selective ejection" (removal of painted egg) and 1 case of "unselective ejection" (rejection error; sensu Moksnes et al., 1991).
Clutch sizes were similar at deserted experimental and control nests (Table 1). Clutch sizes statistically differed (~0.4 eggs) between experimental and control nests that showed "acceptance" (i.e., neither desertion nor ejection response; Table 1).
Clutch characteristics
Clutch size in our NZ population (4.63 ±0.49, n = 41) did not differ significantly from UK source population (4.72 ±0.74, n = 3438; t = 1.23, df = 47, p = 0.23; Fig. 1). Egg length (mm) in NZ population (20.3 ±0.7, n = 41 nests) was similar to the UK population (20.6 ±0.9, n = 100 nests; t = 1.91, df = 139, p = 0.16). Similarly, egg width (mm) in NZ (14.8 ±0.4, n = 41 nests) did not differ from the UK source population (14.8 ±0.7, n = 100 nests; t = 0.00, df = 139, p = 1.00).
Discussion
Egg discrimination
We provide support for the ability of Greenfinches to puncture eject eggs from nests with simulated parasitic eggs. No such ejection events were reported in control nests. Rates of egg ejection did not statistically differ between experimental and control treatments but this can be explained by joint effects of relatively modest sample sizes and species-specifically low rates of egg ejection (both when unselective ejection is included (15%) or excluded (10%) from our sample of experimental nests). Therefore, this statistical non-significance does not imply biological non-significance.
Previous studies with the Greenfinch used hard plastic models that cannot be punctured (Martín-Vivaldi et al., 2002) and all such eggs were accepted (Davies and Brooke, 1989) or some were rejected by desertion or remained in the nest with hosts own eggs destroyed (Moksnes et al., 1991). In contrast, our findings suggest that nest desertion by the Greenfinch might not be a behavioral response to foreign eggs in the nest because it occurred at statistically similar frequencies in both experimental and control nests (see also Kosciuch et al., 2006). Our comparisons of nest desertions between control and experimental nests are based on typical sample sizes in this line of research (n = 20 per treatment) but some other specific results should be interpreted more carefully. Sample sizes are particularly low for comparisons of (1) experimental nests between laying (n = 3) and incubation stages (n = 17) and (2) nest desertion between experimental (n = 3) and control nests during the laying stage (n = 10). Our results suggest that nest desertion was not a specific response to parasitism, because it occurred in both control and experimental nests and at similar rates, but additional data from this and other Greenfinch populations would make for a stronger test of the importance of nest desertion as a potential antiparasitic behavior.
We detected two cases of selective egg ejection of experimentally dyed host eggs and one "unselective ejection" (sensuMoksnes et al., 1991) of own egg (i.e., rejection error), suggesting that Greenfinches are capable of behavioral responses to foreign eggs. Decisions to accept, eject, or desert were not related to clutch size (Table 2). Ejections of manipulated black eggs were most likely an antiparasitism responses because (1) no eggs were ejected at control nests, and (2) hosts at experimental nests specifically targeted the manipulated eggs. In one case a host non-experimental egg was removed by host while the painted egg remained in nest. In a previous study, Boulton and Cassey (2006) used tethered artificial eggs at active nests to identify predators of some European species introduced to NZ, including the Greenfinch. In that study, Greenfinches ejected experimental egg at 1 out of 6 nests providing additional support to our conclusions.
The observed ability of egg ejection and discrimination in the Greenfinch might have evolved due to several selection pressures. (1) Cuckoos in native UK populations may have provided weak but persistent selection pressure for host defenses against interspecific parasites in the Greenfinch. Even though the Greenfinch has traditionally been considered an unsuitable Cuckoo host, some Cuckoo chicks are known to successfully fledge from Greenfinch nests (Seel and Davis, 1981). Although Cuckoos were not introduced to NZ, in some cases antiparasitism adaptations may persist even after the original selection pressures from interspecific parasites ceased (Rothstein, 2001; Hale and Briskie, 2007; Samaš et al., 2011). However, it is important to highlight that the Greenfinch (a) is rare and unsuitable Cuckoo host, (b) has had its NZ population for ~150 years isolated from European Cuckoo, and (c) is not regular host of any native NZ cuckoos (no evidence for NZ cuckoos parasitism on Greenfinches found in literature). Therefore, alternatively or additionally, (2) parasitism by conspecifics may select for an ability to recognize and reject alien eggs (López-de-Hierro and Moreno-Rueda, 2010). Importantly, conspecific brood parasitism is known in the native ranges of Greenfinch (Yom-Tov, 2000). Despite no longer experiencing potential cuckoo parasitism, NZ Greenfinches would likely not escape conspecific parasites and so the conspecific egg rejection hypothesis may provide a viable explanation for persistence of egg rejection behavior (for a similar case of Turdus thrushes see Samaš et al., 2011). We did not find evidence for conspecific parasitism in NZ Greenfinches in literature but detection of such parasitism is much more difficult than the detection of interspecific parasitism (Latif et al., 2006). (3) Also, nest cleaning/ sanitation (sensu Moskát et al., 2003b; Guigueno and Sealy, 2009) might have contributed to the ejection of the dark-painted foreign eggs in our study. However, this hypothesis can explain only non-specific removal of debris (i.e., non-egg-shaped objects) from nests (Ortega and Cruz, 1988; Moskát et al., 2003b; Guigueno and Sealy, 2009) but does not explain specific ability to reject dissimilar eggs (and egg shaped objects) in birds in general (Grim, 2005).
This study emphasized the importance of including control nests for assessing a role of nest desertion in antiparasitic defenses (see also Kosciuch et al., 2006). Our study population showed relatively high rates of desertion of experimentally parasitized nests (20%, 4 out of 20) in comparison to data from source European population (0%, n = 12; Table 3 in Davies and Brooke, 1989). In turn, behavior of the NZ Greenfinch at control nests revealed the same rates of desertion (20%, 4 out of 20) as those at experimental nests. Therefore, nest desertion may not be a specific response to experimental parasitism in our study species.
Clutch characteristics
We did not find significantly different clutch sizes or egg sizes between NZ and UK Greenfinch populations. These results are in contrast with previous studies that showed significantly smaller clutches (Niethammer, 1970; MacMillan, 1985; Evans et al., 2005) and eggs (Cassey et al., 2005) in introduced NZ passerine populations, including Greenfinch. Differences between previous and our work may be caused by different samples used for comparisons.
Conclusions
Importantly, we found that Greenfinches were able to eject foreign eggs although they also may make recognition errors in egg discrimination. That this was observed in a population allopatric from Cuckoos for a long time, and because Greenfinches are rarely parasitized by Cuckoos in their native ranges, suggests that conspecific parasitism, instead of interspecific parasitism, may be responsible for the functional benefit of egg discrimination by Greenfinches (see also Grim et al., 2011; Samaš et al., 2011).
We are grateful to D. Hanley, A. Moksnes, E. Røskaft and an anonymous referee for their helpful comments. This study was supported by Human Frontier Science Program awards (RGY69/07 to TG, PC, and MEH and RGY86/12 to TG and MEH). Our research followed local and institutional guidelines for the treatment of research animals. PC is an ARC Future Fellow.
Antonov A, Stokke BG, Moksnes A, Kleven O, Honza M, Røskaft E. 2006. Eggshell strength of an obligate brood parasite: a test of the puncture resistance hypothesis. Behav Ecol Sociobiol, 60:11–18.
Boulton RL, Cassey P. 2006. An inexpensive method for identifying predators of passerine nests using tethered artificial eggs. New Zeal J Ecol, 30:377–385.
Briskie JV, Mackintosh M. 2004. Hatching failure increases with severity of population bottlenecks in birds. Proc Natl Acad Sci USA, 101:558–561.
Cassey P, Blackburn TM, Evans KL. 2005. Changes in egg size of exotic passerines introduced to New Zealand. Notornis, 53:243–246.
Congdon NM, Briskie JV. 2010. Effect of population bottlenecks on the egg morphology of introduced birds in New Zealand. Ibis, 152:136–144.
Cramp S, Simmons KEL, Perrins CM. 1994. The Birds of the Western Palearctic, Vol. 8: Crows to Finches. Oxford University Press, Oxford.
Davies NB, Brooke ML. 1989. An experimental study of coevolution between the cuckoo, Cuculus canorus, and its hosts. I. Host egg discrimination. J Anim Ecol, 58:225–236.
Evans KL, Duncan RP, Blackburn TM, Crick HQP. 2005. Investigating geographic variation in clutch size using a natural experiment. Funct Ecol, 19:616–624.
Grim T, Samaš P, Moskát C, Kleven O, Honza M, Moksnes A, Røskaft E, Stokke BG. 2011. Constraints on host choice: why do parasitic birds rarely exploit some common potential hosts? J Anim Ecol, 80:508–518.
Grim T. 2005. Mimicry vs. similarity: which resemblances between brood parasites and their hosts are mimetic and which are not? Biol J Linn Soc, 84:69–78.
Grim T. 2006. Cuckoo growth performance in parasitized and unused hosts: not only host size matters. Behav Ecol Sociobiol, 60:716–723.
Grim T. 2007. Equal rights for chick brood parasites. Ann Zool Fennici, 44:1–7.
Guigueno MF, Sealy SG. 2009. Nest sanitation plays a role in egg burial by yellow warblers. Ethology, 115:247–256.
Hale K, Briskie JV. 2007. Response of introduced European birds in New Zealand to experimental brood parasitism. J Avian Biol, 38:198–204.
Hauber ME, Moskát C, Bán M. 2006. Experimental shift in hosts' acceptance threshold of inaccurate-mimic brood parasite eggs. Biol Lett, 2:177–180.
Hauber ME. 2003. Interspecific brood parasitism and the evolution of host clutch sizes. Evol Ecol Res, 5:559–570.
Honza M, Procházka P, Stokke BG, Moksnes A, Røskaft E, Čapek M, Mrlík V. 2004. Are blackcaps current winners in the evolutionary struggle against the common cuckoo? J Ethol, 22:175–180.
Johnson DH. 2002. The importance of replication in wildlife research. J Wildl Manage, 66:919–932.
Kelly CD. 2006. Replicating empirical research in behavioral ecology: how and why it should be done but rarely ever is. Q Rev Biol, 81:221–236.
Kilner RM, Langmore NE. 2011. Cuckoos versus hosts in insects and birds: adaptations, counter-adaptations and outcomes. Biol Rev, 86:836–852.
Kosciuch KL, Parker TH, Sandercock BK. 2006. Nest desertion by a cowbird host: an antiparasite behavior or a response to egg loss? Behav Ecol, 17:917–924.
Lahti DC. 2005. Evolution of bird eggs in the absence of cuckoo parasitism. Proc Natl Acad Sci USA, 102:18057–18062.
Latif QS, Grenier JL, Heath SH, Ballard G, Hauber ME. 2006. First evidence of conspecific brood parasitism and egg ejection in song sparrows, with comments on methods sufficient to document these behaviors. Condor, 108:452–458.
López-de-Hierro MDG, Moreno-Rueda G. 2010. Egg-spot pattern rather than egg colour affects conspecific egg rejection in the house sparrow (Passer domesticus). Behav Ecol Sociobiol, 64:317–324.
Lyon BE, Eadie JMA. 2008. Conspecific brood parasitism in birds: a life-history perspective. Annu Rev Ecol Evol Syst, 39:343–363.
MacMillan BWH. 1985. Breeding of Greenfinches in Hawke's Bay, New Zealand. Notornis, 32:85–93.
Martin TE. 1996. Life history evolution in tropical and south temperate birds: what do we really know? J Avian Biol, 27:263 –272.
Martín-Gálvez D, Soler M, Soler JJ, Martín-Vivaldi M, Palomino JJ. 2005. Food acquisition by common cuckoo chicks in rufous bush robin nests and the advantage of eviction behaviour. Anim Behav, 70:1313–1321.
Martín-Vivaldi M, Soler M, Møller AP. 2002. Unrealistically high costs of rejecting artificial model eggs in cuckoo Cuculus canorus hosts. J Avian Biol, 33:295–301.
Merilä J, Björklund M, Baker AJ. 1996. The successful founder: genetics of introduced Carduelis chloris (greenfinch) populations in New Zealand. Heredity, 77:410–422.
Moksnes A, Røskaft E, Braa AT, Korsnes L, Lampe HM, Pedersen HC. 1991. Behavioural responses of potential hosts towards artificial cuckoo eggs and dummies. Behaviour, 116:64–89.
Moksnes A, Røskaft E. 1992. Responses of some rare cuckoo hosts to mimetic model cuckoo eggs and to foreign conspecific eggs. Ornis Scand, 23:17–23.
Moksnes A, Røskaft E, Solli MM. 1994. Documenting puncture ejection of parasitic eggs by chaffinches Fringilla coelebs and blackcaps Sylvia atricapilla. Fauna Norv Ser C, 17:115–118.
Moskát C, Karcza Z, Csörgö T. 2003a. Egg rejection in European blackbirds (Turdus merula): the effect of mimicry. Ornis Fenn, 80:86–91.
Moskát C, Székely T, Kisbenedek T, Karcza Z, Bártol I. 2003b. The importance of nest cleaning in egg rejection behaviour of great reed warblers Acrocephalus arundinaceus. J Avian Biol, 34:16–19.
Niethammer G. 1970. Clutch sizes of introduced European Passeriformes in New Zealand. Notornis, 17:214–222.
Ortega CP, Cruz A. 1988. Mechanisms of egg acceptance by marsh-dwelling blackbirds. Condor, 90:349–358.
Petrie M, Pinxteen R, Eens M. 2009. Moorhens have an internal representation of their own eggs. Naturwissenschaften, 96:405–407.
Procházka P, Honza M. 2004. Egg discrimination in the yellowhammer. Condor, 106:405–410.
Robinson RA. 2005. BirdFacts: profiles of birds occurring in Britain and Ireland (v1.24, June 2009). BTO Research Report 407, BTO, Thetford. . Accessed 1 December 2012.
Røskaft E, Moksnes A, Stokke BG, Moskát C, Honza M. 2002. The spatial habitat structure hypothesis of host populations explains the pattern of rejection behavior in hosts and parasitic adaptations in cuckoos. Behav Ecol, 13:163–168.
Rothstein SI. 2001. Relic behaviours, coevolution and the retention versus loss of host defences after episodes of avian brood parasitism. Anim Behav, 61:95–107.
Ruxton GD. 2006. The unequal variance t-test is an underused alternative to Student's t-test and the Mann-Whitney U test. Behav Ecol, 17:688–690.
Samaš P, Grim T, Hauber ME, Cassey P, Weidinger, Evans KL. 2013. Ecological predictors of reduced avian reproductive investment in the southern hemisphere. Ecography, doi: .
Samaš P, Hauber ME, Cassey P, Grim T. 2011. Repeatability of foreign egg rejection: testing the assumptions of co-evolutionary theory. Ethology, 117:606–619.
SAS Institute Inc. 2009. JMP version 8.0.1. SAS Institute Inc., Cary, NC.
Seel DC, Davis PRK. 1981. Cuckoos reared by unusual hosts in Britain. Bird Study, 28:242–243.
Servedio MR, Hauber ME. 2006. To eject or to abandon? Life history traits of hosts and parasites interact to influence the fitness payoffs of alternative anti-parasite strategies. J Evol Biol, 19:1585–1594.
Soler JJ, Martínez JG, Soler M, Møller AP. 2001. Life history of magpie populations sympatric or allopatric with the brood parasitic great spotted cuckoo. Ecology, 82:1621–1631.
Soler JJ, Martín-Gálvez D, Martínez JG, Soler M, Canestrari D, Abad-Gómez JM, Møller AP. 2011. Evolution of tolerance by magpies to brood parasitism by great spotted cuckoos. Proc R Soc Lond B, 278:2047–2052.
Stokke BG, Hafstad I, Rudolfsen G, Bargain B, Beier J, Campas DB, Dyrcz A, Honza M, Leisler B, Pap PL. Patapavicius R, Prochazka P, Schulze-Hagen K, Thomas R, Moksnes A, Møller AP, Røskaft E, Soler M. 2007. Host density predicts presence of cuckoo parasitism in reed warblers. Oikos, 116:913–922
Svensson EI, Råberg L. 2010. Resistance and tolerance in animal enemy–victim coevolution. Trends Ecol Evol, 25:267–274.
Thomson GM. 1922. The Naturalisation of Plants and Animals in New Zealand. Cambridge University Press, Cambridge.
Weidinger K. 2010. Foraging behaviour of nest predators at open-cup nests of woodland passerines. J Ornithol, 151:729–735.
White DJ, Ho L, de los Santos G, Godoy I. 2007. An experimental test of preferences for nest contents in an obligate brood parasite, Molothrus ater. Behav Ecol, 18:922–928.
Yom-Tov Y. 2000. An updated list and some comments on the occurrence of intraspecific nest parasitism in birds. Ibis, 143:133–143.
Table
1.
Clutch size (mean ±S.E.) of New Zealand introduced Greenfinches according to their behaviour at experimental and control nests. Differences within the experimental nest category were tested with Kruskal-Wallis test, all other tests were Welsch's t-tests for unequal variances (as recommended by Ruxton, 2006). Sample sizes slightly differ from those in Table 2 because of missing data for one experimental and one control nest.
Table
2.
Overview of New Zealand introduced Greenfinch responses to experimental parasitism (1 host own egg painted black) and control nest treatments. In all, we recorded 3 ejections, 2 cases of "selective ejection" (removal of painted egg) and 1 case of "unselective ejection" (rejection error; sensu Moksnes et al., 1991).

 DownLoad:
DownLoad:





 Email Alerts
Email Alerts RSS Feeds
RSS Feeds