Jesús M. AVILÉS, Deseada PAREJO. 2012: Intra-clutch variation in egg appearance assessed by human vision does not relate to rejection of parasite eggs in Iberian Azure-winged Magpies. Avian Research, 3(4): 302-311. DOI: 10.5122/cbirds.2012.0034
Citation:
Jesús M. AVILÉS, Deseada PAREJO. 2012: Intra-clutch variation in egg appearance assessed by human vision does not relate to rejection of parasite eggs in Iberian Azure-winged Magpies. Avian Research, 3(4): 302-311. DOI: 10.5122/cbirds.2012.0034
Jesús M. AVILÉS, Deseada PAREJO. 2012: Intra-clutch variation in egg appearance assessed by human vision does not relate to rejection of parasite eggs in Iberian Azure-winged Magpies. Avian Research, 3(4): 302-311. DOI: 10.5122/cbirds.2012.0034
Citation:
Jesús M. AVILÉS, Deseada PAREJO. 2012: Intra-clutch variation in egg appearance assessed by human vision does not relate to rejection of parasite eggs in Iberian Azure-winged Magpies. Avian Research, 3(4): 302-311. DOI: 10.5122/cbirds.2012.0034
Estación Experimental de Zonas Áridas, CSIC, Departamento de Ecología Funcional y Evolutiva, Ctra. de Sacramento s/n, 04120, La Cañada de San Urbano, Almería, Spain
The coevolutionary arms race between cuckoos and their hosts predicts that low variation in egg features within a host clutch would facilitate discrimination of mimetic parasite eggs. Here, we experimentally examine this prediction by using artificial and natural parasite eggs showing contrasting level of matching with host eggs. We quantified, based on human assessment, intra-clutch variation in egg appearance and egg discrimination in the Iberian Azure-winged Magpie (Cyanopica cyanus), a presumed former host of the Great Spotted Cuckoo (Clamator glandarius). Azure-winged Magpies rejected parasitic eggs in relation to their degree of dissimilarity with own eggs: Great Spotted Cuckoo model eggs were relatively more often rejected (73.7%) than real Great Spotted Cuckoo eggs (44.4%) and the less contrasting conspecific eggs (35.5%). Contrary to our predictions, we found that, irrespective of mimicry level of parasitic eggs, intra-clutch variation in host egg appearance did not differ significantly between rejecters and acceptors. We found, however, that individuals with higher variation in egg-size were almost significantly more prone to be rejecters than individuals showing lower variation in egg size. Our results do not support the hypothesis that the extent of intraclutch variation in egg discrimination varied with parasite egg mimicry in this particular system, and add to previous findings suggesting that perhaps an increase in intra-clutch variation in egg appearance, rather than a decrease, might be advantageous when discriminating against non-mimetic Great Spotted Cuckoo eggs.
Avian brood parasitism studies have provided us with some of the best examples of co-evolved adaptations (Rothstein, 1990; Davies, 2000). Obligate brood parasites receive parental care from unrelated individuals (hosts) of other species. Successful parasitism often reduces host reproductive output, which favors the evolution of host defenses, leading to selection for more sophisticated trickeries by the parasite to overcome host defenses (Rothstein, 1990). This escalation of adaptations and counter-adaptations by both sides leads to a coevolutionary "arms race" between parasites and hosts (Davies, 2000).
As matching between parasite and host eggs improves, hosts are expected to reduce variation in the appearance of their eggs within their clutches. A low variation in egg appearance within host clutches would facilitate host's recognition of parasite eggs (e.g. Victoria, 1972; Davies and Brooke, 1989b). In addition, a high homogeneity in appearance within clutches would lead to a higher variation in egg appearance among clutches of different host individuals (i.e. inter-clutch variation). Hence, a higher inter-clutch variation in egg appearance will prevent the evolution of a good match for the parasite (Soler and Møller, 1996). Passerine species frequently parasitized by the Common Cuckoo (Cuculus canorus) have evolved a lower degree of intraclutch variation in egg appearance, and a higher degree of inter-clutch variation than those that have not experienced such interactions (e.g. Øien et al., 1995; Soler and Møller, 1996; Stokke et al., 2002; Avilés and Møller, 2003). Recent findings with three Australian hosts of the Pallid Cuckoo (Cuculus pallidus), however, cast doubts on the generality of these processes as intra-clutch variation did not vary between populations with different exposure to parasitism (Landstrom et al., 2011). Intra-specific evidence has so far focused on reporting a link between intra-clutch variation in host egg appearance and probability of rejection of artificial eggs, and, is even more controversial. While some studies have shown that low intra-clutch variation in host egg appearance may favor parasite egg discrimination (Stokke et al., 1999; Soler et al., 2000; Moskát et al., 2008), others did not find support for the parasite recognition function of such egg traits (Karcza et al., 2003; Avilés et al., 2004; Honza et al., 2004; Stokke et al., 2004; Lovászi and Moskát, 2004; Polacikova et al., 2007; Cherry et al., 2007; Landstrom et al., 2010). This variability in results may reflect differences between host species in rejection abilities as the role of intra-clutch variation must be presumably low in good host rejecters as compared to moderate ones (Stokke et al 2007). However, still disparate results might arise due to age-related rejection and egg appearance (e.g. Lotem et al., 1995), and/or a genetic link between egg appearance and rejection behavior (Moskat et al., 2008), that can only be disentangled by experimental manipulations. Indeed, Moskát et al. (2008), showed strong experimental support for the intra-clutch hypothesis as they reported a reduced discrimination of foreign eggs as a consequence of an experimental increase of egg phenotypes variation in a cuckoo host, the Great Reed Warbler (Acrocephalus arundinaceus).
The Azure-Winged Magpie (Cyanopica cyanus) is a social small corvid with a remarkable disjunct distribution. It occurs widely in the eastern Palaearctic and is also found in a smaller geographical area in Spain and Portugal (Cramp and Perrins, 1994). Parasitism of the Azure-winged Magpie by the Common Cuckoo has become very frequent in some parts of the eastern Palearctic (Nakamura et al., 1998). In the Iberian Peninsula two obligate brood parasite species, namely the Great Spotted Cuckoo (Clamator glandarius) and the Common Cuckoo, might potentially exploit the Azurewinged Magpie as host since they are largely sympatric in many areas (Cramp and Perrins, 1994). Although evidence of Common Cuckoo parasitism in Iberia is restricted to one locality in Central Spain (Alonso et al. 1991), evidence coming from egg collections kept in museums suggests that Great Spotted Cuckoo parasitism was widespread at the end of the 19th century because at least 11 cases of parasitized clutches by the Great Spotted Cuckoo were reported in different localities of Spain and Portugal (Avilés, 2004). Indeed, previous studies have shown that Azure-winged Magpies have remarkable discrimination abilities, readily rejecting Great Spotted Cuckoo models and eggs (Arias de Reyna and Hidalgo, 1982; Arias de Reyna, 1998; Avilés, 2004). Also, experiments with adult cuckoo mounts have shown that Azure-winged Magpies defended their nests more aggressively (i.e. higher intensity and shorter latency to an aggressive response) towards Great Spotted Cuckoo and predator dummies than towards nonharmful control species (Avilés and Parejo, 2006).
Here we experimentally parasitized Azure-winged Magpie nests with one of three types of parasite eggs gradually differing in matching from host eggs. In agreement with the arms race hypothesis we predicted: 1) that those individuals with a lower within-clutch variation in egg appearance should be more prone to reject parasite eggs; and, 2) that the role of low intraclutch variation in parasite egg discrimination increased with greater mimicry between parasite and host eggs.
Methods
Study area
The field study was carried out in the surroundings of Guadiloba reservoir near Cáceres (37°18'N, 3°11'W), southern Spain during March–June 2003–2004. The study area approximately comprised 400 ha of a wooded plot of holm oak trees (Quercus ilex) (termed dehesa). Azure-winged Magpie nests were mostly found in holm oak trees.
Study system
The Azure-winged Magpie is a flexible cooperative breeder with occurrence of helper individuals assisting the dominant pair at different stages of the breeding cycle (Valencia et al., 2003). Azure-winged Magpies usually lay five or six eggs (range 2–8) and most first clutches in the study area are found in April and the first week of May (J.M. Avilés, unpublished). Only females incubate the eggs, starting some days after the first egg is laid (Komeda et al., 1987). Egg coloration and maculation varies among females from pale cream or olive-cream backgrounds, rarely with bluish tinge, to pale yellowish-brown or brown-olive, with sparse small brown or olive-brown spots and greyish blotches, often concentrated at the broad end (Cramp and Perrins 1994; Fig. 1). Individual females have a consistent egg type, and based on human assessment their intra-clutch variation is in general very low (Fig. 1). Great Spotted Cuckoo eggs are elliptical to sub-elliptical, with blunt ends, smooth and fairly glossy, pale green-blue, thickly spotted, light brown or red brown color (Cramp and Perrins 1994; Fig. 1) and they are not good mimics to Azure-winged Magpie eggs according to the human eye (Fig. 1). Also spectrophotometric analyses of Azurewinged Magpie clutches parasitized by Great Spotted Cuckoos saved in European egg collections revealed a poor level of mimicry between cuckoo and Azurewinged Magpie host eggs (Soler et al., 2003).
Figure
1.
Variation in egg appearance within clutches of Azurewinged Magpie (a, b). Representative differences in appearance between Azure-winged Magpie eggs (five top eggs) and experimental Azure-winged Magpie egg in the bottom (c); Representative appearance of model Great Spotted Cuckoo egg (d), and real Great Spotted Cuckoo eggs (e) used in the experiments. Mean dimensions of experimental eggs were: model Great Spotted Cuckoo eggs = 32.5 mm×25.1 mm; real Great Spotted Cuckoo eggs = 31.4 mm×23.8 mm, Azure-winged Magpie eggs = 26.7 mm×20.0 mm.
We searched for nests at the beginning of the breeding seasons 2003–2004. 76 nests were found at different stages of building and were numbered and mapped. The nests were visited daily and each egg was numbered with waterproof ink in consecutive order as laid. We did not detect evidence of intraspecific parasitism since only one egg ever appeared on a single day. Eight Azure-winged Magpie nests were predated before responses to artificial parasitism could be assessed. The remaining 68 nests were randomly assigned to one of the following groups: Experimentally parasitized with Great Spotted Cuckoo model eggs (n = 19), experimentally parasitized with real Great Spotted Cuckoo eggs (n = 18) or experimentally parasitized with conspecific eggs (n = 31). Size dimensions of eggs in each of these groups are reported in the Fig. 1. The three experimental groups were not equally distributed between the two years of study. Experiments with Great Spotted Cuckoo model eggs were performed in 2003, while response to real Great Spotted Cuckoo eggs was assessed in 2004 only. However, parasitism with conspecific eggs was performed in 2003 (n = 14) and 2004 (n = 17). Since the study was undertaken for 2 seasons, and birds were not banded, there is a possibility that the same individuals have been included in two experiments.
Great Spotted Cuckoo model eggs were quail (Coturnix japonica) eggs painted with acrylic paint to mimic the color and spotted pattern of real cuckoo eggs in the area (Avilés, 2004, Fig. 1). Real Great Spotted Cuckoo eggs were collected from deserted Magpie (Pica pica) nests. We did not quantify variation between cuckoo eggs in our population, but they all showed a considerably degree of mismatching with host eggs according to the human eye (Fig. 1). Conspecific eggs were collected from recently abandoned nests in the study area. We avoided using conspecific eggs that were abandoned more than a week before artificial parasitism because eggs may fade in color due to storage (Moreno et al., 2011). Experimental eggs were not reused and no host eggs were removed during experimental parasitism to simulate Great Spotted Cuckoo laying tactics and conspecific parasitism (Avilés, 2004).
Dimensions of Great Spotted Cuckoo models were within the range size and mass of real Great Spotted Cuckoo eggs measured in the area, but slightly larger than the average (Fig. 1). In addition, although models were painted to match the color and pattern of a typical cuckoo egg in the area, it should be noted that to the human eye real Great Spotted Cuckoo eggs achieved a better matching of host eggs than our models (Fig. 1). Hence, matching in egg size and appearance between host and parasite eggs greatly varied among the three experimental treatments ranging from poor (cuckoo models) to good (conspecific eggs), with real Great Spotted Cuckoo eggs being at an intermediate stage (Fig. 1).
The time interval between laying of the first host egg and the beginning of the experiment varied. 47.0% of nests (n = 32) were artificially parasitised during host egg laying period, and the remaining 53.0% (n = 36) were parasitised 2–3 days after completion of the clutch. However, no breeding phase effect on rejection is expected since response to artificial parasitism in the same Azure-winged Magpie population did not vary between laying and incubation (Avilés, 2004). In no case did artificial parasitism cause host eggs to be removed the subsequent days. All nests were checked every second day and response to parasitism was assessed six days from artificial parasitism as rejection (eggs ejected or nest deserted) or acceptance of parasitic eggs (eggs still warm and incubated in nest).
Measurements of intra-clutch variation
Experimental eggs were always added to Azure-winged Magpie nests after the laying of the fifth egg, which is about one day in advance to clutch completion. Thus, in all experimental nests, hosts were able to assess intraclutch variation of their entire clutch before experiencing artificial parasitism. At that time we obtained two different measures of intra-clutch variation in host eggs based on human assessment: one accounting for variation in color and spottiness estimated from photographs, and the second yielding variation in egg size. Because we took photos one day before clutch completion we were not able to assess the total intra-clutch variation (at least one egg laid afterwards). However, the last egg laid is not different in appearance from the others in Azure-winged Magpies (J.M. Avilés, pers. obs.) making it unlikely that the appearance of the last egg may affect our results. Host clutches were photographed in a standardized way with a Nikon Coolpix 990 digital camera. Clutches were numbered and photographed on a standard Kodak neutral-grey card. Intra-clutch variation in egg color and marking was later assessed on a 1–5 scale (1 = no variation, 5 = all the eggs differ from each other) (Øien et al., 1995) by five experienced ornithologists that did not know the fate of the nest in advance and that were unaware of the hypothesis being tested. Consistency of the five scores was significant (repeatability = 0.51, F19, 80 = 6.28, p < 0.0001), and one mean score value per clutch as an estimation of intraclutch variation in egg color and marking was then calculated. In addition, length and width of all eggs were taken using a caliper with a precision of 0.01 mm. Egg volume was later estimated from the formula V = 0.515LB2 (Hoyt, 1979), and the standard deviation in egg volume within a clutch was used in further analyses as a measure of intra-clutch variation in egg size.
We are aware that our method of estimating variation in egg appearance was based upon human vision and hence does not account for UV information, which birds might perceive (e.g. Cherry and Bennett, 2001; Avilés and Møller, 2003; Avilés et al., 2006). However, human assessments of intra-clutch variation have the advantage of incorporating patterns of speckling of the eggs, which are not easily tractable by using spectrophotometry.
Statistical methods
We performed logistic regression models with the GENMOD procedure of SAS (SAS Institute, 1996) to test for the association between the occurrence of rejection (rejection vs. acceptance of parasitism) and the two measures of intra-clutch variation and the type of experimental egg introduced into the Azure-winged Magpie nests. Probability of rejection was modeled as a binomial response variable (1 = rejection, 0 = acceptance) using a logistic link function. To account for the fact that the three experimental groups were not equally distributed in the two years (see data collection) we introduced the experimental treatment nested in the study year and the study year as a fixed effect in the model.
Previously, the existence of age-related rejection and egg color features for some cuckoo hosts have been reported (e.g. Lotem et al., 1992). To account for this possibility we tested for the associations between rejection of artificial eggs and clutch size that is assumed an age-dependent feature among passerine birds (e.g. Lack, 1968). Furthermore, we tested for the possible association between variation in egg traits and host clutch size by using Pearson correlations (GLM procedure in SAS). Starting models contained all the main effects plus all possible interactions. Model selection was carried out by removing, one by one, the effects that were the furthest to statistical significance, starting with the highest-order interactions down to the main effects.
Results
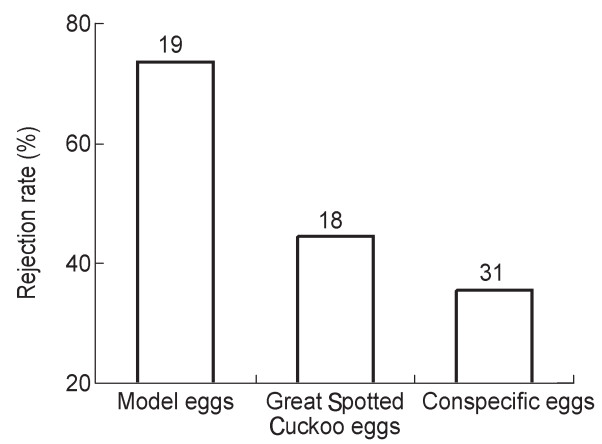
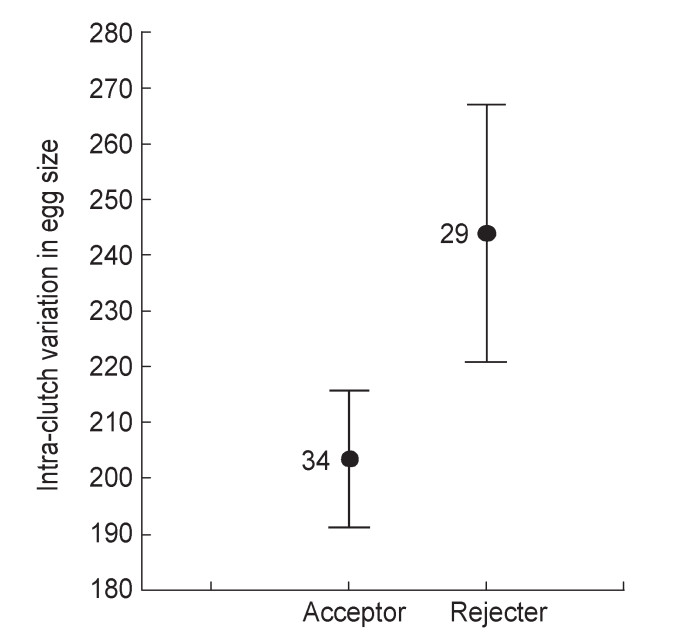
Proneness to rejection by Azure-winged Magpies increased with degree of dissimilarity between introduced experimental eggs and host eggs (Table 1): Great Spotted Cuckoo model eggs were more frequently rejected (73.68%) than real Great Spotted Cuckoo eggs (44.44%) and than the more mimetic conspecific eggs (35.5%, Fig. 2). In addition, we found that intra-clutch variation in egg size, but not in egg color and marking, almost significantly related to the probability of accepting or rejecting parasitic eggs by Azure-winged Magpies (Table 1). Individuals with higher variation in egg-size were more prone, although not significantly (p = 0.07), to be rejecters than individuals showing lower variation in egg size (Fig. 3).
Table
1.
Relationship between rejection of parasitic eggs and the type of parasitic egg (Great Spotted Cuckoo model, Great Spotted Cuckoo and conspecific eggs), intra-clutch variation in egg color and marking (ITCM), intra-clutch variation in egg size (ITS) and year
χ2
df
p
Egg type×ITCM×ITS
2.19
2, 47
0.33
ITCM×ITS
2.51
1, 49
0.11
Egg type×ITCM
4.27
2, 50
0.12
Egg type×ITS
2.90
2, 52
0.23
ITCM
2.82
1, 54
0.09
ITS
3.21
1, 58
0.07
Egg type
7.40
2, 58
0.025
Study year
0.13
1, 58
0.72
Tests performed with generalized linear models with occurrence of rejection (rejection vs. acceptance of parasitism) as a binomial dependent variable and the two measures of intra-clutch variation, the type of experimental egg nested in the study year and the study year as independent variables. Model selection started from the complete model. All effects and interactions are shown irrespective of level of significance.
Figure
2.
Egg rejection in relation to level of mimicry between the parasite and Azure-winged Magpie eggs. Rejection rate increased with dissimilarity between parasite and host eggs after accounting for unequal distribution of experimental eggs in the two years (χ2 = 7.40, df = 2, 58, p = 0.025).
Figure
3.
Mean (±S.E.) intra-clutch variation in egg size (i.e., S.D. in volume within cluth) in relation to the response to artificial parasitism (rejection vs. acceptance). Sample sizes are shown close to the mean values.
We found no support for the existence of age-related responses to parasitism by Azure-winged Magpies. First, the probability of Azure-winged Magpies rejecting model eggs was not significantly associated with clutch size (mean = 5.87, S.D. = 0.18, n = 35 in acceptors vs. mean = 6.09, S.E. = 0.15, n = 32 in rejecters, χ2 = 1.00, df = 1, 65, p = 0.32). Furthermore, neither intraclutch variation in egg color and marking (rp = -0.14, p = 0.28, n = 60), nor intra-clutch variation in egg size (rp = -0.11, p = 0.39, n = 60) were significantly associated to Azure-winged Magpie clutch size.
Discussion
Obligate avian brood parasites may greatly reduce host fitness and then actively select for the evolution of antiparasite defensive mechanisms in their hosts. In the coevolutionary arms race between cuckoos and their hosts, discrimination and rejection of parasite eggs by hosts is a main defensive mechanism reducing parasite prevalence and favoring the evolution of parasite egg mimicry (Davies, 2000). Discrimination abilities are then expected to be a function of degree of matching between parasite and host eggs (e.g. Davies, 2000). In addition, it has been suggested that a reduction in intra-clutch variation in egg appearance may help hosts in discriminating against cuckoo eggs highly resembling their ones (e.g. Victoria, 1972; Davies and Brooke, 1989a). As expected, Azure-winged Magpies rejected parasitic eggs in relation to its degree of dissimilarity with their own eggs (Fig. 2, Table 1): nonmimetic Great Spotted Cuckoo model eggs were more frequently rejected than real cuckoo eggs; and these were more frequently rejected than the conspecific eggs. This tendency has been shown by several previous studies on other host species (e.g. Davies and Brooke, 1988, 1989a; Soler and Møller, 1990; Moksnes et al., 1993; Lotem et al., 1995; Lathi and Lathi, 2002; Procházka and Honza, 2003, 2004; Cassey et al., 2008; Avilés et al., 2010), and was suggested to be the factor selecting for mimicry in eggs of the parasite. The trend that we have found would be predicted by either variation in color and marking or variation in size of parasite eggs across the three experimental groups. Therefore, it is difficult to discriminate among degree of matching in color and marking, and in size as the possible factors eliciting rejection of parasitic eggs in the Azure-winged Magpie.
Our results do not support the first prediction concerning the relationship between intra-clutch variation in egg appearance and discrimination of parasite eggs. We found a non-significant trend for variation in egg size, but not in color or marking, to be important (but p = 0.07) explaining rejection behavior against parasite eggs in the Azure-winged Magpie. Those individuals with a higher intra-clutch variation in egg size within their clutches showed a non-significant trend to reject more frequently than those individuals with a lower intra-clutch variation in egg size (Fig. 3). In addition, our results did not fit the second prediction concerning an increasing role of low intra-clutch variation in parasite discrimination as mimicry between the parasite and the host eggs increases.
To date a large number of studies using Common Cuckoos or Great Spotted Cuckoos as the target parasite species have assessed the adaptive role of intra-clutch variation in terms of parasite egg discrimination at the intra-specific level (see introduction). Support for this hypothesis came from Stokke et al. (1999), which found the expected negative relationship between intra-clutch variation and rejection probability. Also Moskat et al. (2008) found that when you experimentally altered intra-clutch variation (making it higher than controls), rejection rate decreased in great reed warbler hosts. However, the more prevalent result found in other studies was a no significant relationship between intraclutch variation in egg appearance and parasite egg rejection (Karcza et al., 2003; Avilés et al., 2004; Honza et al., 2004; Stokke et al., 2004; Lovászi and Moskát, 2004; Polacikova et al., 2007; Landstrom et al., 2010). Discrepancies between studies may be due to the different level of recognition abilities for the tested hosts as moderate rejecters might differently rely on intra-clutch variation in egg appearance than good rejecters (Stokke et al., 2007). Also, Polacikova et al. (2007) showed that it may be features at the egg blunt pole rather that those estimated for the whole egg that are important.
Expectation from the intra-clutch variation hypothesis for the hosts of the Great Spotted Cuckoo should differ from that for hosts of the Common Cuckoo because the former parasite has eggs that do not mimic host eggs (Soler et al., 2003), which would reduce the adaptive value of a low intra-clutch variation in terms of egg discrimination (Victoria, 1972; Davies and Brooke, 1989). Then we may expect intra-clutch variation to be unimportant in terms of discrimination of parasite eggs among Great Spotted Cuckoo hosts. Indeed, results from the three only studies analyzing the hypothesis with hosts of the Great Spotted Cuckoo do not support this prediction. In fact, two studies have found some support for a positive relationship between intra-clutch variation and discrimination abilities (Avilés et al., 2004; this study), whereas the third study found a negative relationship (Soler et al., 2000). This fact may suggest that hosts of Great Spotted Cuckoo, in which parasite-host matching is rather poor (Soler et al., 2003), are using discrimination rules different than the ones assumed for Common Cuckoo hosts when discarding parasite eggs. For instance, it could be advantageous for the hosts to produce eggs with signatures because a foreign egg not bearing such a signature could be easily discriminated. Alternatively, rejection and intra-clutch variation in egg appearance may be age dependent traits in the Azurewinged Magpie. Since egg discrimination by hosts is likely based on a learning mechanism (Victoria, 1972; Lotem et al., 1992), the detected pattern should be a consequence of the greater recognition abilities of parasitic eggs by older Azure-winged Magpies not linked to their egg appearance. However, this explanation seems to be unlikely for this system, because clutch size, that is assumed to be an age-related trait in passerines (e.g. Lack, 1968), was unrelated to rejection rate and to intraclutch variation in egg features (this study).
Alternatively, our results could be related to other selective pressures acting on azure-winged magpie egg appearance. As well as interspecific brood parasitism, intraspecific brood parasitism and nest predation are factors potentially shaping egg appearance in birds (see review in Underwood and Sealy, 2002). Victoria (1972) predicted that just as in species that are affected by interspecific brood parasitism, a reduction in intra-clutch variability and an increase in inter-clutch variability might increase the chance of discrimination of conspecific eggs when the probability of intraspecific brood parasitism is high. Indeed, Azure-winged Magpies have the potential of discarding conspecific eggs which suggests that egg rejection in this species may have evolved from intra-specific parasitism (Avilés, 2004). However, in long-term studies of the species in Spain (reviewed in Avilés, 2004; Valencia et al., 2004) in which eggs were daily numbered, no evidence of intraspecific brood parasitism was found. Likewise, preliminary studies of parentage of the Azure-winged Magpie revealed no evidence of intraspecific brood parasitism (J.G. Martínez, pers. commun.). On the other hand, nest predation might differently affect individual hosts as a function of the variation in the appearance of their eggs. However, in only 10% of the studies where eggs were artificially placed in nests did changes in egg coloration explain changes in egg predation (Underwood and Sealy, 2002). In addition, we failed to find any link between intraclutch variation in egg color and matching (One-way ANOVA, F1, 28 = 0.16, p = 0.69) and egg size (One-way ANOVA, F1, 28 = 0.11, p = 0.70) and probability of predation while performing our experiments, suggesting a minor role for nest predation on our results.
In conclusion, our results support the view that Azure-winged Magpies have remarkable cognitive abilities for discriminating against parasite eggs. We found that Azure-winged Magpies rejected parasitic eggs in relation to its degree of dissimilarity with their own eggs. We also found that Azure-winged Magpies that rejected foreign eggs showed an almost significant higher variation in egg size than acceptors, and that the role of intra-clutch variation in parasite egg discrimination does not change with mimicry. The most likely explanation for our results is that Azure-winged Magpie egg coloration may favor recognition of both intra- or inter-specific parasitism because variation in egg appearance was consistently linked to a higher recognition ability of parasite eggs. Considering together ours and previous findings in magpie hosts parasitized by great spotted cuckoos suggesting that a large variation in egg appearance within host clutches may favor parasite egg discrimination, future studies should focus on the study of the plausible discrimination mechanisms leading to such a relationship.
Acknowledgements
We thank Carlos de la Cruz for advice on how to search for Azure-winged Magpie nests. Pablo Cidoncha provided invaluable help in the field. We are grateful to the Dirección General de Medio Ambiente de la Junta de Extremadura which provided us with permissions, and to the owners and guards of Casa Urraca and Suerte de Santa Maria who provided us with facilities during field work. We thank two anonymous referees and the guest editors, Eivin Røskaft and Bård Gunnar Stokke that provided constructive comments on the manuscript. This work was performed without financial support.
Alonso JA, Muñoz-Pulido R, Bautista LM, Alonso JC. 1991. Nest-site selection and nesting success in the Azure-winged Magpie in Central Spain. Bird Study, 38:45–51.
Arias de Reyna L, Hidalgo SJ. 1982. An investigation into eggacceptance by azure-winged magpies and host-recognition by great spotted cuckoo chicks. Anim Behav, 30:819–823.
Arias de Reyna L. 1998. Coevolution of the great spotted cuckoo and its hosts. In: Rothstein SI, Robinson SK (eds) Parasitic Birds and Their Hosts: Studies in Coevolution. Oxford University Press, New York, pp 129–142.
Avilés JM, Møller AP. 2003. Meadow pipits (Anthus pratensis) egg appearance in cuckoo (Cuculus canorus) sympatric and allopatric populations. Biol J Linn Soc, 79:543–549.
Avilés JM, Parejo D. 2006. Nest defense by Iberian azure-winged magpies (Cyanopica cyanus): do they recognize the threat of brood parasitism? Ethol, Ecol Evol, 18:321–333.
Avilés JM, Soler JJ, Pérez-Contreras T, Soler M, Møller AP. 2006. Ultraviolet reflectance of great spotted cuckoo eggs and egg discrimination by magpies. Behav Ecol, 17:310–314.
Avilés JM, Soler JJ, Soler M, Møller AP. 2004. Egg appearance and rejection behaviour in magpies. Anim Behav, 67:951–958.
Avilés JM, Vikan JR, Fossoy F, Antonov A, Moksnes A, Røskaft E, Stokke BG. 2010. Avian color perception predicts behavioral responses to experimental brood parasitism in chaffinches. J Evol Biol, 23:293–301.
Avilés JM. 2004. Egg rejection by Iberian azure-winged magpies in absence of brood parasitism. J Avian Biol, 35:295–299.
Avilés JM. 2008. Egg colour mimicry in the common cuckoo Cuculus canorus as revealed by modelling host retinal function. Proc R Soc B, 275:2345–2352.
Brooke ML, Davies NB. 1988. Egg mimicry by cuckoos Cuculus canorus in relation to discrimination by hosts. Nature, 335:630–632.
Cassey P, Grim T, Honza M, Hauber ME. 2008. The modeling of avian visual perception predicts behavioural rejection responses to foreign egg colours. Biol Lett, 4:515–517.
Cherry MI, Bennett ATD, Moskat C. 2007 Host intra-clutch variation, cuckoo egg matching and egg rejection by great reed warblers. Naturwissenschaften, 94:441–447.
Cherry MI, Bennett ATD. 2001. Egg colour matching in an African cuckoo, as revealed by ultraviolet-visible reflectance spectrophotometry. Proc R Soc Lond B, 268:565–571.
Cramp S, Perrins CM. 1994. Handbook of the Birds of Europe, the Middle East and North Africa: the Birds of the Western Palearctic, vol. Ⅷ. Oxford University Press, Oxford.
Davies NB, Brooke ML. 1988. Cuckoos versus reed warblers: adaptations and counteradaptations. Anim Behav, 36:262–284.
Davies NB, Brooke ML. 1989a. An experimental study of coevolution between the cuckoo, Cuculus canorus, and its hosts. Ⅰ. Host egg discrimination. J Anim Ecol, 58:207–224.
Davies NB, Brooke ML. 1989b. An experimental study of coevolution between the cuckoo, Cuculus canorus, and its hosts. Ⅱ. Host egg marking, chick discrimination and general discussion. J Anim Ecol, 58:225–236.
Davies NB. 2000. Cuckoos, Cowbirds and Other Cheats. T. and A. D. Poyser, London.
Honza M, Procházka P, Stokke BG, Moksnes A, Røskaft E, Čapek Jr M, Mrlík V. 2004. Are blackcaps current winners in the coevolutionary struggle against the common cuckoo? J Ethol, 22:175–180.
Hoyt D. 1979. Practical methods of estimating volume and fresh weight of bird eggs. Auk, 96:73–77.
Karcza Z, Moskat C, Cherry MI, Kisbenedek T. 2003. Experimental manipulation of intraclutch variation in the great reed warbler shows no effect on rejection of parasitic eggs. Ethology, 109:15–22.
Komeda S, Yamagishi S, Fujioda M. 1987. Cooperative breeding in azure-winged magpies, Cyanopica cyana, living in region of heavy snowfall. Condor, 89:835–841.
Lack D. 1968. Ecological Adaptations for Breeding in Birds. Methuen, London.
Landstrom MT, Heinsohn R, Langmore NE. 2010. Clutch variation and egg rejection in three hosts of the pallid cuckoo, Cuculus pallidus. Behaviour, 147:19–36.
Landstrom MT, Heinsohn R, Langmore NE. 2011. Does clutch variability differ between populations of cuckoo hosts in relation to the rate of parasitism? Anim Behav, 81:307–312.
Lathi DC, Lathi AR. 2002. How precise is egg discrimination in weaverbirds? Anim Behav, 63:1135–1142.
Lotem A, Nakamura H, Zahavi A. 1992. Rejection of cuckoo eggs in relation to host age: a possible evolutionary equilibrium. Behav Ecol, 3:128–132.
Lotem A, Nakamura H, Zahavi A. 1995. Constraints on egg discrimination and cuckoo-host co-evolution. Anim Behav, 49:1185–1209.
Lovászi P, Moskát C. 2004. Break-down of arms race between the red-backed shrike (Lanius collurio) and common cuckoo (Cuculus canorus). Behaviour, 141:245–262.
Marchetti K. 2000. Egg rejection in a passerine bird: size does matter. Anim Behav, 59:877–883.
Moksnes A, Røskaft E, Korsnes L. 1993. Rejection of Cuckoo (Cuculus canorus) eggs by Meadow pipits (Anthus pratensis). Behav Ecol, 4:120–127.
Moreno J, Lobato E, Morales J. 2011. Eggshell blue-green colouration fades immediately after oviposition: a cautionary note about measuring natural egg colours. Ornis Fenn, 88:51–56.
Moskát C, Avilés JM, Ban M, Hargitai R, Zolei A. 2008. Experimental support for the use of egg uniformity in parasite egg discrimination by cuckoo hosts. Behav Ecol Sociobiol, 62:1885–1890.
Moskát C, Honza M. 2002. European Cuckoo Cuculus canorus parasitism and host's rejection behaviour in a heavily parasitised Great Reed Warbler Acrocephalus arundinaceus population. Ibis, 144:614–622.
Nakamura H, Kubota S, Suzuki R. 1998. Coevolution between the common cuckoo and its major hosts in Japan. In: Rothstein SI, Robinson SK (eds) Parasitic Birds and Their Hosts: Studies in Coevolution. Oxford University Press, New York, pp 94–112.
Øien IJ, Moksnes A, Røskaft E. 1995. Evolution of variation in egg color and marking pattern in European passerines: adaptations in a coevolutionary arms race with the cuckoo, Cuculus canorus. Behav Ecol, 6:166–171.
Payne RB. 1997. Avian brood parasitism. In: Clayton DH, Moore J (eds) Host Parasite Coevolution. General Principles and Avian Models. Oxford University Press, Oxford, pp 338–369.
Polaciková L, Honza M, Procházka P, Topercer J, Stokke BG. 2007. Colour characteristics of the blunt pole: cues for recognition of parasitic eggs as revealed by reflectance spectrophotometry. Anim Behav, 74:419–427.
Procházca P, Honza M. 2003. Do common whitethroats Sylvia communis discriminate against alien eggs? J Ornithol, 144:354–363.
Procházca P, Honza M. 2004. Egg discrimination in the yellowhammer. Condor, 106:405–409.
Rothstein SI. 1975. Evolutionary rates and host defenses against avian brood parasitism. Am Nat, 109:161–176.
Rothstein SI. 1990. A model system for coevolution: avian brood parasitism. Annu Rev Ecol Syst, 21:481–508.
Rutila J, Latja R, Koskela K. 2002. The common cuckoo Cuculus canorus and its cavity nesting host, the redstart Phoenicurus phoenicurus: a peculiar cuckoo-host system? J Avian Biol, 33:414–419.
SAS Institute. 1996. SAS/STAT software changes and enhancements through release 6.11. SAS Institute. North Carolina.
Soler JJ, Avilés JM, Soler M, Møller AP. 2003. Evolution of host egg mimicry in a brood parasite, the great spotted cuckoo. Biol J Linn Soc, 79:551–563
Soler JJ, Møller AP. 1996. A comparative analysis of the evolution of variation in appearance of eggs of European passerines in relation to brood parasitism. Behav Ecol, 7:89–94.
Soler JJ, Soler M, Møller AP. 2000. Host recognition of parasite eggs and the physical appearance of host eggs: the Magpie and its brood parasite the Great Spotted Cuckoo. Etología, 8:9–16.
Soler M, Møller AP. 1990. Duration of sympatry and coevolution between the great spotted cuckoo and its magpie host. Nature, 343:748–750.
Spottiswoode CN, Stevens M. 2010. Visual modeling shows that avian host parents use multiple visual cues in rejecting parasitic eggs. PNAS, 107:8672–8676.
Stoddard MC, Stevens M. 2010. Pattern mimicry of host eggs by the common cuckoo, as seen through a bird's eye. Proc R Soc B, 277:1387–1393.
Stokke BG, Moksnes A, Røskaft E, Rudolfsen G, Honza M. 1999. Rejection of artificial cuckoo (Cuculus canorus) eggs in relation to variation in egg appearance among reed warblers (Acrocephalus scirpaceus). Proc R Soc Lond B, 266:1483–1488.
Stokke BG, Moksnes A, Røskaft E. 2002. Obligate brood parasites as selective agents for evolution of egg appearance in passerine birds. Evolution, 56:199–205.
Stokke BG, Rudolfsen G, Moksnes A, Røskaft E. 2004. Rejection of conspecific eggs in chaffinches: the effect of age and clutch characteristics. Ethology, 110:459–470.
Stokke BG, Takasu F, Moksnes A, Roskaft E. 2007. The importance of clutch characteristics and learning for antiparasite adaptations in hosts of avian brood parasites. Evolution, 61:2212–2228.
Underwood TJ, Sealy SG. 2002. Adaptive significance of egg coloration. In: Deeming DC (ed) Avian Incubation. Oxford University Press, Oxford, pp 280–298.
Valencia J, Cruz C, Carranza J, Solís E. 2004. No brood parasitism by the great spotted cuckoo Clamator glandarius on the azure-winged magpie Cyanopica cyanus. Bird Study, 52:204–209.
Valencia J, Cruz C, González B. 2003. Flexible helping behaviour in the azure-winged magpie. Ethology, 109:1–15.
Victoria JK. 1972. Clutch characteristics and egg discriminate ability of the African village weaverbird Ploceus cucullatus. Ibis, 114:367–376.
Table
1.
Relationship between rejection of parasitic eggs and the type of parasitic egg (Great Spotted Cuckoo model, Great Spotted Cuckoo and conspecific eggs), intra-clutch variation in egg color and marking (ITCM), intra-clutch variation in egg size (ITS) and year
χ2
df
p
Egg type×ITCM×ITS
2.19
2, 47
0.33
ITCM×ITS
2.51
1, 49
0.11
Egg type×ITCM
4.27
2, 50
0.12
Egg type×ITS
2.90
2, 52
0.23
ITCM
2.82
1, 54
0.09
ITS
3.21
1, 58
0.07
Egg type
7.40
2, 58
0.025
Study year
0.13
1, 58
0.72
Tests performed with generalized linear models with occurrence of rejection (rejection vs. acceptance of parasitism) as a binomial dependent variable and the two measures of intra-clutch variation, the type of experimental egg nested in the study year and the study year as independent variables. Model selection started from the complete model. All effects and interactions are shown irrespective of level of significance.


 DownLoad:
DownLoad:





 Email Alerts
Email Alerts RSS Feeds
RSS Feeds