V. Gokula. 2011: Breeding biology of the Spot-billed Pelican (Pelecanus philippensis) in Karaivetti Bird Sanctuary, Tamil Nadu, India. Avian Research, 2(2): 101-108. DOI: 10.5122/cbirds.2011.0013
Citation:
V. Gokula. 2011: Breeding biology of the Spot-billed Pelican (Pelecanus philippensis) in Karaivetti Bird Sanctuary, Tamil Nadu, India. Avian Research, 2(2): 101-108. DOI: 10.5122/cbirds.2011.0013
V. Gokula. 2011: Breeding biology of the Spot-billed Pelican (Pelecanus philippensis) in Karaivetti Bird Sanctuary, Tamil Nadu, India. Avian Research, 2(2): 101-108. DOI: 10.5122/cbirds.2011.0013
Citation:
V. Gokula. 2011: Breeding biology of the Spot-billed Pelican (Pelecanus philippensis) in Karaivetti Bird Sanctuary, Tamil Nadu, India. Avian Research, 2(2): 101-108. DOI: 10.5122/cbirds.2011.0013
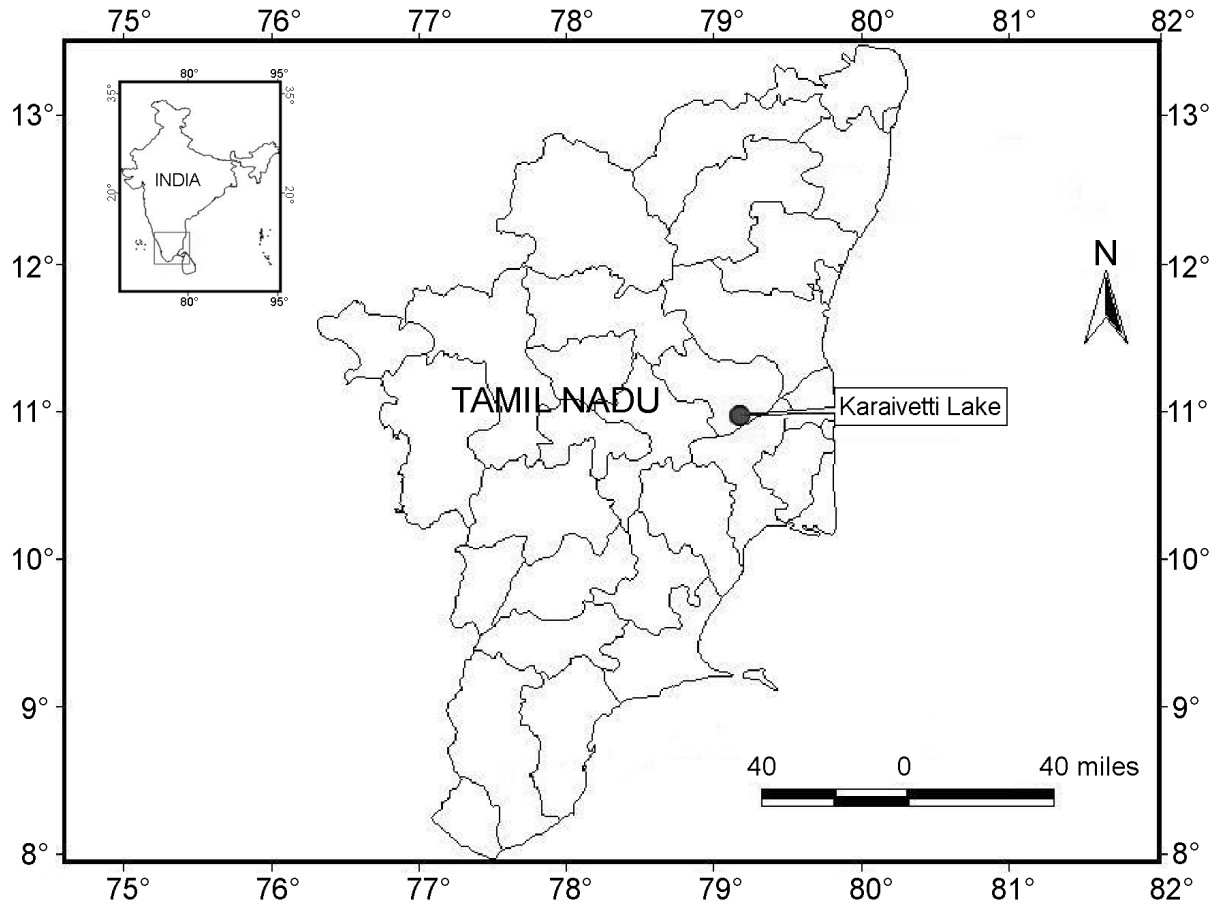
The breeding biology of the Spot-billed Pelican (Pelecanus philippensis) was investigated from September 2007 to September 2010 in Karaivetti Lake, Tamil Nadu, India. The Spot-billed Pelican breeds between November and April at the Karaivetti Lake. It also breeds roughly during the same period at other breeding sites in Tamil Nadu, except for the Vedanthangal and Karikili bird sanctuaries, where pelicans arrives much earlier and their breeding activities start and end to some extent ahead of the other sites, i.e., from September to March. In essence, the breeding season starts after the onset of the northeast monsoon in Tamil Nadu. A few days after their arrival, pelicans perform courtship display, form pairs, start nest construction and lay eggs. Courtship display is not as attractive and impressive as in other waterbirds. The clutch size varies from two to three eggs and the incubation period from 25 to 36 days. The fledging period varies from a minimum of 90 to a maximum of 102 days. The nest success rate of the Spot-billed Pelican at Karaivetti was 90.28% during the study period.
Avian eggs have strong calcareous shells serving several functions such as mechanical protection, gas exchange and providing calcium and phosphorus for the developing embryo. Eggshell strength is related allometrically to egg size (Ar et al., 1979) but specific selection regimes have shaped unusually strong eggs in some avian taxa such as penguins (Boersma et al., 2004) and holenesting ducks (Mallory and Weatherhead, 1990) that lay their eggs on rough nesting substrates. Another example is the Marsh Wren (Cistothorus palustris) in which stronger than expected eggshells withstand habitual and frequent egg destruction by conspecifics (Picman et al., 1996; Picman and Honza, 2002). Another major group of birds well known for increased eggshell strength is the obligate brood parasites. Phylogenetically distant brood parasitic birds such as cuckoos, cowbirds and honeyguides all lay structurally stronger eggs than their closest non-parasitic relatives (Spaw and Rohwer, 1987; Picman, 1989; Picman and Pribil, 1997; Spottiswoode and Colebrook-Robjent, 2007; Spottiswoode, 2010). This indicates that eggshell strength may be an adaptation to their brood parasitic life-style. Eggshell strength is achieved by increased eggshell thickness, rounder shape, higher eggshell density and enhanced micro-hardness; the contribution of these mechanisms may differ among brood parasitic taxa (Picman, 1989; Brooker and Brooker, 1991; Picman and Pribil, 1997; Igic et al., 2011). Eggshells contain up to 98% calcium and many birds have been shown to be calcium-limited during laying (reviewed in Reynolds et al., 2004). Since brood parasites lay many more eggs than nonparasitic birds (Payne, 1977), the maintenance of thick and/or unusually dense eggshells may be particularly costly, also indirectly pointing to their putative adaptive function in the face of potential calcium limitation. However, cuckoos eat many host eggs and may hence replenish their calcium supply this way (Moksnes et al., 2000). Furthermore, as recently discovered, hostparasite covariation in eggshell strength suggests that eggshell strength may be a coevolutionarily evolved trait between brood parasites and their hosts just as egg mimicry (Spottiswoode, 2010). However, it is still unclear in what ways increased eggshell strength increases fitness of brood parasites. Two major hypotheses attempt to explain the adaptive significance of the unusual eggshell strength in brood parasites. Here we review the evidence in favour of each of these and also consider two additional alternative explanations. Finally, we identify several major areas of future research that will potentially elucidate evolutionary hypotheses regarding strong eggshells in brood parasites.
Laying damage hypothesis
The "laying damage hypothesis", originally proposed for the Common Cuckoo (Cuculus canorus) (Lack, 1968), states that the increased structural strength of parasitic eggs serves to decrease the risk of its damage by collision with host eggs during the typically rapid laying of brood parasites (Chance, 1940; Sealy et al., 1995; Moksnes et al., 2000). Rapid laying is beneficial to brood parasites because some hosts may either inflict serious injury on the laying parasite by direct attack (Molnar, 1944; Davies, 2000), or be more likely to reject the parasite egg if the parasite is detected at the nest (Davies and Brooke, 1988; Moksnes et al., 1993). Thus, the need to lay in haste together with evidence that some cuckoo hosts are cavity nesters or build domed nests where the parasite cannot enter to lay directly suggests that cuckoo eggs might indeed benefit from their strong eggshells during the process of laying (Wyllie, 1981). A necessary condition assumed by the laying damage hypothesis is that parasitic eggs are projected into the host nest from so large a distance that there is a real risk of egg damage (Spaw and Rohwer, 1987). Several videos of laying cuckoos clearly show that the females lay from some distance rather than sitting on the nest. Because there are normally some (typically weakershelled) host eggs in the nest at the time of parasitism, we would expect to see frequent damage to host eggs indicative of collision, i.e., dents and/or cracks (Spaw and Rohwer, 1987). The laying damage hypothesis is apparently relevant for Clamator cuckoos, which typically have the opportunity to lay directly in host nests, but nevertheless lay their eggs by 'shooting' them against the host clutch from a considerable distance (e.g. 15 cm in one documented case, Gaston 1976). Observational and experimental data have shown that egg damage of Magpie (Pica pica) eggs in the form of dents or cracks typically occur in association with laying events by the Great Spotted Cuckoo (Clamator glandarius) (Soler et al., 1997; Soler and Martínez, 2000). These cuckoos parasitize relatively large and/or cooperatively breeding host species, hence the need for rapid egg laying is particularly important because of the risk of hosts seriously injuring the parasite if caught at the nest (Gaston, 1976; Soler et al., 1997; Soler, 1990; Davies, 2000; Soler and Martínez, 2000). However, Soler et al. (1997) provided experimental evidence that laying damages caused this way may be an adaptive strategy since, upon hatching, the parasitic chick is left with fewer host chick competitors to start with and has higher probability of survival. Clamator cuckoos do not evict host chicks, but monopolize host parental effort by outcompeting host chicks via exaggerated begging displays (Redondo, 1993, 1999). Whichever of the two mechanisms may provide a stronger selective advantage, both are beneficial to the brood parasite and are thus non-exclusive explanations. There is less support for the laying damage hypothesis in cowbirds Molothrus spp. and those in the genus Cuculus, since these parasites typically have the opportunity to sit on the host nest to lay, and there is no evidence of collision damage of host eggs (Spaw and Rohwer, 1987; Rothstein and Robinson, 1998; Davies, 2000; Antonov et al., 2006a, 2008a, 2008b). However, Common Cuckoos and other large-sized cuckoos do in many cases use hosts that are considerably smaller than themselves, building small nests that the cuckoo cannot sit on properly. Hence, videos have shown that cuckoos in such cases have to lay its egg from distance and thereby increasing the risk of own or host egg damage. The importance of the laying damage hypothesis is therefore still largely unexplored, and future studies should aim to study parasite laying behavior and damage to host and parasite eggs by close monitoring and video recordings.
Puncture resistance hypothesis
The "puncture resistance hypothesis", also originally proposed for cuckoos, but later elaborated for cowbirds, states that eggshell strength represents an adaptation which renders rejection by hosts difficult and/or costly, thereby increasing the probability that parasitic eggs are accepted (Swynnerton, 1918; Spaw and Rohwer, 1987; Rohwer and Spaw, 1988; Krüger, 2011). Due to body and/or bill-size constraints, the mode of rejection of parasitic eggs varies among host species and can be either grasp ejection, puncture ejection or desertion (Davies and Brooke, 1989a; Moksnes et al., 1991a). The most cost-efficient mode of rejection is grasp ejection, i.e. selective removal of the parasitic egg from the nest by the host while saving the rest of the clutch (Rothstein, 1975; Lorenzana and Sealy, 2001). However, many small host species are unable to grasp the parasitic egg in their mandibles due to constraints imposed from small bills or particular bill shapes (Rothstein 1975; Moksnes et al., 1991a; but see Underwood and Sealy, 2006). Such hosts have to first puncture the parasitic egg to be able to take it in the bill and carry it away from the nest (Rohwer and Spaw, 1988; Moksnes et al., 1991a, 1994). However, puncture ejection can be costly because hosts often damage some of their own eggs while trying to puncture the parasitic egg and such egg losses are termed ejection costs (Moksnes et al., 1994). In addition, puncturing the parasitic egg may require considerable effort and time, and pecking behavior is also incompatible with incubation (Martín-Vivaldi et al., 2002; Soler et al., 2002; Antonov et al., 2008b). Puncture ejections may bring additional costs via spillage of egg content in the nest, which may cause further damage to the remaining host eggs by causing them glue to each other and/or to nest lining (Rothstein, 1975; Antonov, 2010), as well as increase the risk of infestation of the nest by ants (Clark and Robertson, 1981) and microorganisms (Soler et al., 2012a). However, experiments have shown that such additional costs of puncture ejection are not substantial in at least two hosts of the common cuckoo (Antonov et al., 2006a, 2009). Given that the vast majority of both cuckoo and cowbird hosts are small passerines (Davies, 2000), rejection costs in terms of own egg damage are likely to be a sufficiently important factor to drive the evolution of increased eggshell strength in brood parasites (Spaw and Rohwer, 1987; Rohwer and Spaw, 1988). Unfortunately, reliable data on the extent to which different host species suffer rejection costs are very scarce because most experiments used impenetrable artificial eggs (overestimating costs, e.g., Moksnes et al., 1991a) or real eggs with eggshells less strong than real parasitic eggs (underestimating costs, e.g., Rothstein, 1975; Martín-Vivaldi et al., 2002; Stokke et al., 2002). However, the few experiments with real parasitic eggs in a few hosts of both cowbirds (e.g., Rohwer et al., 1989; Røskaft et al., 1993; Sealy and Neudorf, 1995) and more recently cuckoos (Antonov et al., 2006a, 2008a, 2008b) have shown that small hosts often suffer substantial costs trying to reject real parasitic eggs. The mode of rejection and the extent and magnitude of costs have been shown to depend on host bill-size with larger-billed species rejecting by grasp ejection experiencing no or very few costs and smallerbilled ones suffering progressively greater costs as bills become smaller (Davies and Brooke, 1989a; Moksnes et al., 1991a; Antonov et al., 2006a). However, this relationship is not straightforward because there are documented cases where larger-billed hosts suffer larger costs than much smaller hosts (Underwood and Sealy, 2006).
The most important prerequisite for the puncture resistance hypothesis is that strong-shelled eggs must translate into fitness benefits for the parasite, i.e. be accepted more often than thin-shelled "normal" eggs of the same size. This can theoretically work providing that the host can raise some of its offspring alongside the parasitic one, and the costs of accepting the parasite are less than the costs of abandoning the current breeding attempt and renesting. Thus, it may pay such hosts to 'forcibly' accept the hard-shelled parasitic egg that they cannot selectively remove from the nest (Spaw and Rohwer, 1987; Rohwer and Spaw, 1988). Therefore, rejection costs in terms of the ejection of own eggs in unparasitized nests have been considered in theoretical models trying to explain the spread of rejection genes in host populations, and they can affect the evolution of different types of defence against parasite eggs (Davies and Brooke, 1989b; Moksnes et al., 1991; Lotem and Nakamura, 1998; Røskaft and Moksnes, 1998). Parasitized hosts of cowbirds and Clamator cuckoos may obtain some reproductive success since parasitic chicks of these parasites are raised alongside host offspring ('nonevicting' parasites). Thus, rejection costs may play a role in favouring acceptance over the costly ejection in such host-brood parasite systems whenever the mean number of host eggs damaged during ejection exceeds the equivalent mean number of host fledglings lost to raising a parasite (Rohwer and Spaw, 1988).
Nevertheless, empirical evidence for cowbirds and their hosts shows that the above condition may not be fulfilled even for relatively large host species such as the Northern Oriole (Icterus galbula) (Røskaft et al., 1993). Furthermore, chicks of these non-evicting brood parasites often outcompete host chicks of smaller species so that foster parents raise few or no young of their own (Becking, 1981; Marvil and Cruz, 1989; Soler, 1990; Lorenzana and Sealy, 2001). In addition, hosts of both cowbirds and Clamator cuckoos also experience higher levels of multiple parasitism, which further contributes to the total reproductive failure by the hosts (Friedmann et al., 1977; Soler, 1990). However, there is a twist to the cowbird story as the cowbird chick has been experimentally shown to extract more parental feedings from hosts and grow better when raised together with some host chicks in at least one medium-sized host species (Kilner et al., 2004). Overall, in the majority of these cases, the hosts of non-evicting parasites seem to do better by abandoning the parasitized clutch and renesting, and rejection costs seem of little importance in making hosts accept (Lorenzana and Sealy, 2001; Underwood and Sealy, 2006). Thus, the widespread acceptance of cowbird eggs, or the predominance of nest desertion as the rejection mode by many small hosts, is considered to be better explained by evolutionary lag (Underwood and Sealy, 2006). The latter authors even suggested that the puncture resistance hypothesis should not be relevant for cowbirds, but instead for cuckoos since cuckoo hosts are more prone to commit recognition or rejection errors, hence a more phenotypically plastic response may have been favoured (Underwood and Sealy, 2006). The chicks of 'evicting' brood parasites such as most parasitic cuckoos and honeyguides eliminate all host progeny by eviction or direct killing, monopolizing parental care and reducing the benefits of accepting parasitism to zero (Davies, 2000; Payne, 2005). Thus, in such host-parasite systems, rejection costs have traditionally been considered unlikely to have an evolutionary influence on host responses. Rather, a parasitic egg can only escape rejection as long as it is not detected by the host as foreign (Spaw and Rohwer, 1987), implying that the puncture resistance hypothesis cannot work in evicting brood parasites.
In line with Underwood and Sealy (2006), we argue here that a strong eggshell can still convey fitness benefits even to such virulent parasites as evicting cuckoos, although we refrain from subscribing to the idea that this is necessarily working better in cuckoos than in cowbirds. Host rejection decisions were shown to have a conditional component and are influenced by the perceived risk of actual parasitism, i.e., whether the parasite was detected at the nest (Davies and Brooke, 1989a; Moksnes et al., 1993), or if host and cuckoo populations are sympatric (Lindholm, 2000; Lindholm and Thomas, 2000; Stokke et al., 2008). Such conditional responses are thought to be beneficial because they minimize the risk of recognition errors, i.e., erroneous rejection of odd-looking host eggs in the absence of brood parasitism (Davies et al. 1996; Røskaft et al., 2002). This problem is exacerbated in host-parasite systems in which the parasite has evolved highly mimetic eggs. Assuming this, signal detection modelling indicates that when the probability of being parasitized is low (as is usually the case in most hosts of the common cuckoo at least), an acceptance strategy may be favoured by selection (Davies et al., 1996). The degree of cuckoo egg mimicry is likely associated with a continuum of host motivational states extending to complete rejection. Thus, if the phenotype of the foreign egg is close to the threshold of host cognitive abilities (Rodríguez-Gironés and Lotem, 1999; Stokke et al., 2007b), some small puncture ejector hosts may peck the foreign egg for some time, but cease the effort to complete puncture due to of their low motivation. Such a scenario is especially likely for hosts with costly ejection methods, short breeding seasons and rapidly seasonally declining prospects of successful re-nesting (Moksnes et al., 1993; Lotem and Nakamura, 1998). Since some cuckoo eggs fail to hatch due to infertility/addling (Øien et al., 1998), it may pay such a host to continue with the parasitized clutch under some circumstances. There has been at least one study showing that Reed Warblers may peck non-mimetic model eggs, but sometimes accept them without desertion (Lindholm, 2000). However, since model eggs are impossible to puncture eject, it has been difficult to establish whether and how frequently this ever occurs in nature to be of any evolutionary significance for cuckoohost systems. A series of recent studies on two small hosts of the Common Cuckoo, involving experimental introduction of real cuckoo eggs and manipulations of egg mimicry, found mixed support for the puncture resistance hypothesis (Antonov et al., 2008a, 2008b, 2009). Marsh Warbler (A. palustris) individuals, a major current Common Cuckoo host in Europe (Moksnes and Røskaft, 1995), were able to invariably puncture and eject cuckoo eggs once they were recognized as foreign, no matter how long it took host individuals to complete rejection. Rejection rate of real cuckoo eggs painted non-mimetic did not differ significantly from rejection rate of similarly-sized and coloured, but much weakershelled eggs; also none of the mimetic real cuckoo eggs which were pecked (i.e., recognized) was accepted (Antonov et al., 2008a). Thus, the strong eggshell of the cuckoo egg does not seem to be important in this host species, which has advanced defences against brood parasitism and likewise also in similarly-sized heavily exploited host species. On the other hand, the slightly smaller Olivaceous Warbler (Hippolais pallida) accepted a considerable proportion of real non-mimetic cuckoo eggs, which were otherwise pecked and hence recognized as foreign. In one instance, an Olivaceous Warbler female damaged and ejected two own eggs, yet eventually accepted the cuckoo egg (Antonov et al., 2009). Interestingly, some Olivaceous Warblers were able to successfully puncture eject the cuckoo egg, although such birds were in minority (Antonov et al., 2009). Its average body mass is slightly smaller compared to that of the Marsh Warbler (Cramp, 1998). Thus, the apparently inferior ability of the Olivaceous Warbler to puncture eject may be due to constraints if this species is positioned close to the threshold of physical characteristics necessary to complete puncture ejection. Alternatively, all Olivaceous Warbler individuals may be fully able to accomplish puncture ejection, although the costs of doing so may be higher than in Marsh Warblers. Surprisingly, we recorded a very low frequency of rejection costs in Olivaceous Warblers (Antonov et al., 2009), although this may be precisely because these hosts do not try hard enough in their rejection attempts to avoid them. Olivaceous Warblers are one of the latest arriving breeding passerines in the study area, and thus they have a short breeding season (Antonov et al., 2007a). Clutch size in this species declines markedly with laying date, suggesting that re-nesting attempts may be costly perhaps due to declining food supplies during the dry summer (Antonov et al., 2007a). A high incidence of acceptance of cuckoo eggs and phenotypic plasticity in rejection behavior due to short breeding seasons was also suggested for Meadow Pipits (Anthus pratensis) breeding in mountains at high latitudes (Moksnes and Røskaft, 1987; Moksnes et al., 1993).
An additional factor that may contribute to the observation that egg discrimination may not necessarily be followed by rejection could be the low motivation of birds to reject (Lindholm, 1999). Cuckoo parasitism of Olivaceous Warblers has shown a decline from heavy (ca. 27% in 2001–2003 (Antonov et al., 2007b) to 0% in 2006–2009 (Antonov et al., unpublished data)), perhaps related to an accompanying decline in the size of that local population of Olivaceous Warbler hosts (Stokke et al., 2007a). Nevertheless, cuckoos were still present in their breeding habitat, implying that Olivaceous Warblers must have perceived the risk of parasitism. Since equivalent experiments in marsh warblers were always carried out at unparasitized nests during the same time interval, we do not regard the temporal absence of natural parasitism in the Olivaceous Warbler population as a serious confounding factor in the observed differences in rejection decisions in Olivaceous and Marsh Warblers.
Thus, forced acceptances obviously occur in nature in some small host species of even evicting (highly virulent) brood parasites suggesting that the puncture resistance hypothesis is not irrelevant for such systems, as previously thought (Spaw and Rohwer, 1987; Davies, 2000). Information on the variation in the actual mode of rejection in Common Cuckoo hosts is still scarce, but it seems that most hosts are small puncture ejecting passerines (Moksnes et al., 1991), and forced acceptances may well be found in a number of other species as more studies document host rejection behavior by video-recordings.
Chick vigour hypothesis
The eggshell is, as the primary source of calcium, necessary for the development of embryonic skeleton and muscle tissue (Simkiss, 1961; Blom and Lilja, 2004). The remarkable behavior displayed by the chicks of evicting cuckoos soon after hatching in ejecting all host progeny may necessitate a more advanced development of its skeleton, i.e., a higher degree of bone ossification than should be expected for an altricial hatchling of similar size. Climbing up the nest cup wall while carrying its load would also require well developed leg musculature. Ejection of host eggs or nestlings shortly after hatching is very important for cuckoo chick growth and survival because it may otherwise fail to outcompete host siblings (Rutila et al., 2002; Martin-Galvez et al., 2005; Hauber and Moskat, 2008; Grim et al., 2011). Therefore, it is possible that cuckoo eggshells may be selectively neutral in terms of egg survival, but important as a source of calcium for a more advanced development of the skeleto-muscular system of the cuckoo chick as an adaptation to its eviction behavior. One study attempted to show that Common Cuckoo hatchlings even possess special morphological adaptations to facilitate hatching from a strong-shelled egg (Honza et al., 2001), indicating that at the time of hatching eggshells are still too strong to be broken open by the hatchling. However, that study compared the morphology of newly hatched cuckoo chicks with that of one of its main passerine hosts, the Great Reed Warbler (Acrocephalus arundinaceus), having similarly sized eggs. Since there may be intrinsic differences between cuckoos and passerines in hatchling morphology, comparing a parasite with its distantly related host fails to determine whether cuckoo chicks have evolved special adaptations to facilitate hatching from strong-shelled eggs relative to their nonparasitic ancestor. Further research should compare hatchling morphology of parasitic birds and their nonparasitic closest relatives to address this possibility. A prediction from the chick-vigour hypothesis is that the eggs of non-evicting parasites should be thinner-shelled relative to their size than those of evicting cuckoos. However, this seems not to be the case as the eggs of the non-evicting Clamator cuckoos are thicker-shelled relative to those of evicting Cuculus/Cacomantis/Chrysococcyx species (Becking, 1981; Brooker and Brooker, 1991). Another prediction from the chick vigour hypothesis is that parasites with host races, e.g., like the well-studied Common Cuckoo, should have thicker-shelled eggs in host races exploiting larger hosts. A similar prediction can be made for the laying damage hypothesis, while the opposite would be the case for the puncture resistance hypothesis. Such a comparison is lacking and this possibility remains to be tested. The degree of calcium mobilization from the eggshell by the developing embryo in birds is related to the speed of embryonic development, with the faster growing species consuming less of the eggshell calcium reserves and vice versa in species with slower development, while controlling for body size (Blom and Lilja, 2004). Since cuckoo embryos develop even faster than those of their passerine host species, a substantial amount of chick bone ossification as a result of elevated utilization of the eggshell calcium reserve is unlikely, although direct studies clearly are necessary. In any case, the chick vigour hypothesis is limited in its generality as it addresses only evicting brood parasites, and it fails in its most important prediction.
Anti-bacterial protection hypothesis
At the species level, cuckoos and cowbirds are generalist brood parasites. For instance, at least 221 host species of the Brown-headed Cowbird (Molothrus ater) are known (Friedmann and Kiff, 1985), and more than 125 species are known to have been parasitized by the Common Cuckoo in Europe (Davies 2000). Hence, these parasites inevitably experience different bacterial communities, and it is difficult to evolve maternally transmitted immune factors against all. Soler et al. (2012a) showed that eggshells of Great Spotted Cuckoos had lower bacterial density than their corvid hosts, which are known to encounter many and diverse microorganisms due to their habit of eating carrion. In addition, the density of bacteria inside unhatched and hence unviable eggs was higher for the Magpie (Pica pica) than for the Great Spotted Cuckoo. Parasite eggs will always experience the bacterial community of the host, while most hosts will only experience their own bacterial community. Thus there should be strong selection on parasites to develop antimicrobial defences against the bacteria of hosts, while the reverse would not be the case. A thick eggshell is a mechanical defence against microorganisms (Board et al., 1994), and embryo mortality due to microorganisms is common in chicken (Baggott and GraemeCook, 2002) and in many species of wild birds (Soler et al., 2012b). Extensive studies of eggshell strength in chickens have provided evidence for an optimal egg shape with an optimal size and density of eggshell pores (Solomon et al., 1994). The density and size of eggshell pores that maximizes eggshell strength is not necessarily optimal for prevention of bacterial infection of eggs. Therefore, we suggest that there is potential for a tradeoff between eggshell properties that promote strength and those that prevent infection with microorganisms. Hence, it is possible that the thick and more compacted eggshells of brood parasites function as a generalized mechanical barrier against bacterial contamination that is more efficient than that of their hosts.
A list of the hypotheses for the evolution of thickshelled eggs in brood parasites, the main selective pressures and the predictions are listed in Table 1. To summarize, while there is still uncertainty regarding the question of how strong-shelled eggs confer fitness benefits to brood parasites, based on existing evidence, the most likely and general explanation seems to be the puncture resistance hypothesis. The laying damage hypothesis can account for strong eggshells in some nonevicting parasites such as the Clamator cuckoos, and possibly also in Cuculus cuckoos.
Table
1.
Summary of main selective pressure increasing eggshell thickness and major predictions of the four hypotheses for the evolution of thick eggshells in brood parasites
Laying damage hypothesis
Puncture resistance hypothesis
Chick vigour hypothesis
Anti-bacterial protection hypothesis
Main selective pressure
Damage to host eggs and prevention of damage to parasite eggs
Ejection of parasite eggs made difficult for hosts
Calcium in eggs allows precocial development of skeleto-muscular system in brood parasite chick
Thicker eggshell should provide greater protection against microorganisms
Damage to host eggs
Yes
Yes
Not important for hypothesis
Not important for hypothesis
Distance from laying cuckoo to host eggs
Dropped from distance except in small parasites that utilize hosts of similar size
Not important for hypothesis
Not important for hypothesis
Not important for hypothesis
Damage to host eggs reduces competition with host chicks
Yes
Not important for hypothesis
Not important for hypothesis
Not important for hypothesis
Damage to hole- or domenesting species common
Yes
Not important for hypothesis
Not important for hypothesis
Not important for hypothesis
Shell thickness of evicting and non-evicting parasites
No difference
No difference
Relatively thinner eggs in non-evicting parasites
No difference
Shell thickness in different host races
Relatively thicker eggs in parasite eggs from host races exploiting hosts with stronger eggshells
Relatively thicker eggs in parasite eggs from host races exploiting puncture ejecter species
Relatively thicker eggs in parasite eggs from host races exploiting hosts with larger eggs
Relatively thicker eggs in parasite eggs from host races exploiting larger hosts because larger hosts have more bacteria
Parasite eggshell strength and the evolution of rejection modes
The strength of eggshells seems to be related to ejection behavior, which shows considerable variation among populations and species. As revealed by experiments with model eggs, some hosts of the Common Cuckoo, e.g., the Great Reed Warbler which is relatively large in terms of body mass and bill-size and is readily able to use grasp ejection of cuckoo eggs, always first try to puncture eggs (Lotem et al., 1995; Antonov et al., 2006a; for similar results see also Honza and Moskat, 2008). Comparisons with the evidence from several similar studies on other Sylviidae suggest that puncture ejection is the ancestral mode of rejection in this family (Martín-Vivaldi et al., 2002; Procházka and Honza, 2003; Honza et al., 2004). Obviously grasp ejection is also simultaneously present in the behavioral repertoire of some species physically able to do so. Likewise, both ejection methods are documented in diverse hosts of the Brown-headed Cowbird (Rothstein, 1976, 1977; Sealy and Neudorf, 1995; Sealy, 1996; Rasmussen et al., 2009). Since even attempts at grasp ejections were not recorded in small hosts such as Marsh Warblers and Olivaceous Warblers (Antonov et al., 2008b, 2009), these species do not seem to be physically capable of grasp ejecting a cuckoo egg and can only use puncture ejection as a rejection method. However, the puncture ejection mode may be fixed in some species, i.e., they are unable to change their responses when parasitized with a small model egg that is impossible to puncture but possible to grasp eject. For example, experiments have shown that Common Whitethroats (Sylvia communis), a strong rejecter of non-mimetic eggs (Procházka and Honza, 2003), lack the flexibility to switch to grasp ejection under experimentally manipulated settings in which grasp ejection was made possible. Instead these hosts invariably desert after failing to puncture a small model egg (Antonov et al., unpublished data). Clearly, flexibility in rejection modes across a variety of hosts, differing in their stages of the arms races with their brood parasites merits further investigation.
Video-recordings of actual egg rejection behavior in small host species, unable to grasp eject indicate that desertion as a passive abandonment of parasitized clutches may not be a true response to the presence of a brood parasitic egg in the nest. For example, Olivaceous Warblers appear to reject most naturally laid Common Cuckoo and model eggs by desertion (Antonov et al., 2007b), although video-recordings confirmed that desertions seem to represent failed attempts to puncture the foreign egg (Antonov et al., 2009). This also seems to be the case in Cedar Waxwings (Bombycilla cedrorum) (Rothstein, 1976). Furthermore, Yellow Warblers (Dendroica petechia) reject cowbird eggs by unselective burial or desertion, although some individuals were shown to first peck and even succeeding in puncture ejection in rare cases (Guigueno and Sealy, 2010; S.G Sealy in preparation). Another example is Meadow Pipits that reject cuckoo eggs by true nest abandonment (no pecking observed) in Norway (Moksnes et al., 1993), although some ejections were recorded in UK (Davies and Brooke, 1989a). Because of the widespread use of impenetrable model eggs in egg rejection experiments, it is unclear whether and to what extent desertion is a genuine anti-parasite defence in hosts. Birds can also desert nests in response to disturbance and the sight of predators at the nest, and brood parasites are common nest predators (Wyllie, 1981; Gärtner, 1982; Schulze-Hagen, 1992). Since desertion is present in the behavioral repertoire of hosts outside the context of brood parasitism, desertion is thought of as the most primitive response type to brood parasitism (Hosoi and Rothstein, 2000). Many studies have failed to show that desertion is a specific response to the presence of a foreign egg per se more than clutch reduction caused by brood parasites in association with laying, the sight of the adult parasite at the nest, or disturbance by visiting investigators (e.g., Clark and Robertson, 1981; Moksnes et al., 1993; Hosoi and Rothstein, 2000; Antonov et al., 2006b; Kosciuch et al., 2006). Thus, species rejecting brood parasite eggs by apparent desertion may either 1) be relatively recent hosts and thus more efficient rejection modes such as grasp or puncture ejection have not yet evolved, or 2) desertion is an evolutionary stable response having been established since host body size and/or beak morphology constrains ejection. To conclude, there is a need for comparative analyses to link eggshell strength to interspecific variation in antiparasite behavior among hosts.
Proficiency at egg ejection as an evolved trait modulated by individual experience
Egg ejection behavior shows considerable variation among individuals, and the link between such phenotypically plastic behavior and eggshell strength remains unknown. A video-recording study has recently shown that host individuals within a population show substantial variation in their efficiency at performing puncture ejection after the parasitic egg has been identified (Antonov et al., 2008b). In this study, many Marsh Warblers which are physically capable of completing the puncture ejection process in 1–2 minutes of focused strong pecking delayed ejections for many hours by pecking too weakly to produce a hole in the cuckoo egg (Antonov et al., 2008b). Such seemingly suboptimal behavior was hypothesized to be due to the existence of a learning component, i.e., host individuals facing the parasitic egg for the first time need to learn to adjust pecking strength before being able to make a hole in the eggshell, while individuals having already been parasitized and rejected cuckoos eggs, eject more efficiently in subsequent parasitism events. By extension, the frequency of ejection costs experienced by a host individual may also be negatively related to prior exposure of hosts to parasitism. However, we hypothesize that the mechanics underlying ejection behavior may have a genetic component enabling it to evolve in coevolutionary arms races, similar to the ability to recognize foreign eggs. Since virtually all studies in avian host-brood parasite systems have only focused on the cognitive component of egg discrimination and the final output (rejection/acceptance), the evolution of ejection proficiency remains virtually unknown. If the complex of behavioral steps leading to ejection, and efficiency of this process evolve, then we should see consistently different rejection costs among host species not strictly explicable by the physical abilities of these species to perform ejection. Egg damage during ejection is documented in various hosts of both cuckoos and cowbirds. While the incidence and magnitude of costs depend on host bill characteristics (Rohwer and Spaw, 1988; Moksnes et al., 1991a, 1991b), this is certainly not the only factor explaining ejection costs. For example, ejecting Warbling Vireos (Vireo gilvus) suffer much less egg damage than the largerbodied and larger-billed Baltimore Oriole (Icterus galbula) (Underwood and Sealy, 2006). Since the Warbling Vireo is one of the smallest-billed ejectors among cowbird hosts, these authors invoked evolutionary lag as the most likely explanation for why ejection is not more ubiquitous among larger-billed hosts (Underwood and Sealy, 2006). Indeed, rejection by ejection seems to be much more frequent among the hosts of cuckoos as these systems are considered evolutionarily much older than cowbirds and their hosts (Hosoi and Rothstein, 2000). In a comparative analysis we should expect a negative relationship between the overall rejection rate and the frequency of costs in different host species, after controlling for host grasp index. Furthermore, as the rejection modes of the different hosts become known, rejection rates should be positively related to the relative frequency of grasp to puncture ejection within ejecting species, again after controlling for grasp index. The incidence of weak pecking in at least one host of the Common Cuckoo was shown to be positively related to foreign egg mimicry (Antonov et al., 2008b). Thus, weak pecking was proposed to be adaptive when the host is unsure whether a foreign egg is a cuckoo egg as this would minimize the risk of misdirected pecks at own eggs once host motivation to reject has increased (Antonov et al., 2008b). To conclude, there is a need for investigating the role of phenotypic plasticity in ejection behavior and how this relates to variation in eggshell strength. Perspectives and avenues for future research This review has emphasised a number of apparently contradictory findings in the study of eggshell strength in brood parasitic birds. Experimental parasitism of hosts of brood parasites has proven an invaluable research tool for better understanding this model system of coevolution (Rothstein, 1990). However, we must accept that this area of research may have suffered from problems of experimental procedures because hosts behave differently when responding to artificial eggs and real cuckoo eggs (Martín-Vivaldi et al., 2002). This comment is not to diminish the efforts of previous research, but to further our understanding of the behavior of interactions between hosts and parasites. Thus, our understanding of the behavioral mechanisms of ejection of parasite eggs is in need of a re-assessment. A largely unexplored field of research is to what extent grasp and puncture ejection are both available as options in the behavioral repertoire of the different host species. Preliminary data showed that Bramblings (Fringilla montifringilla), Chaffinches (F. coelebs) and other strongly rejecting host species of the Common Cuckoo are very good puncture rejecters of parasitic eggs, although their specialization as puncture ejecters may have gone so far that they have lost flexibility in the ability to switch to grasp ejection in a context-dependent fashion (Moksnes et al., 1993; Antonov et al., unpublished). Thus, a comparative analysis of the flexibility in egg rejection modes in different host species may be illuminating. Given the seemingly suboptimal rejection performance documented within a single population of at least one common cuckoo host, it is worth exploring further whether the efficiency at ejection has a learnt component. Future experiments involving repeated measures of rejection performance of known-age individuals would cast light on this problem (Honza et al., 2007).
A related problem is whether there is a heritable component to egg discrimination, i.e., the sequence of actions needed to complete puncture ejection successfully once a foreign egg has been detected. If so, and given that egg rejection behavior is often costly (see above), we can predict that such behavior can decay faster than egg recognition abilities in host populations freed from parasitism, e.g., in hosts introduced to novel geographic areas lacking parasites, or when initially parasitized hosts historically become isolated from their parasite for natural reasons. Thus, such host populations may have retained egg recognition, but show deficiencies in the process of rejection, leading them to accept non-mimetic parasitic eggs even if they recognize them as foreign. Only future experiments coupled with video-recordings can cast light on this interesting possibility.
Finally, an exciting future area of research concerns exploration of whether different host races or species of brood parasites differ in eggshell strength. A necessary pre-condition to favor increased eggshell strength in some parasitic lineages is that their host species should be puncture ejectors, and that puncture ejection should be difficult to accomplish and/or costly. Given that birds are calcium-limited, and that eggshell strength must be traded against the ability of the cuckoo chick to hatch (Honza et al., 2001), it would not pay cuckoos to increase eggshell strength in host species readily able to reject parasitic eggs. Thus, cuckoo eggs are predicted to be thinner-shelled in accepting, grasp ejecting and efficient puncture ejecting host species, and strongershelled in the smallest puncture ejectors since only in the latter case may increase eggshell strength lead to 'forced' acceptance (Antonov et al., 2009). A recent study provided some support for these predictions. Spottiswoode (2010) showed a positive relationship between egg rejection frequencies and eggshell strength in five cuckoo host races in Great Britain, although this relationship was mainly caused by the thin-shelled eggs of the cuckoo host race parasitizing the Dunnock (Prunella modularis). Dunnocks are universal acceptors of non-mimetic cuckoo eggs (Davies and Brooke, 1989a; Moksnes et al., 1991a), and thus next to no selection on eggshell strength in the respective cuckoo host race is expected from the parasite. Yet, Dunnocks are exceptional among Common Cuckoo hosts, the vast majority of which show some rejection of naturally laid cuckoo eggs (Davies, 2000). The fact that cuckoo eggshell thickness has decreased in this host species indicates indirectly that maintenance of this trait is costly and given no selection for it exerted by the host, it will not persist. More recently, Igic et al. (2011) compared cuckoo eggshell strength in three European cuckoo host races, by using several advanced mechanical measures including micro-structural strength analysis, failed to find significant differences among host races. Thus, a larger sample of host species and their host races of brood parasites, including a wider spectrum of hosts in terms of their egg rejection frequencies and modes of rejection, are needed to rigorously test these findings. Interestingly, Spottiswoode (2010) also found significant covariation in eggshell strength of Diederic's Cuckoo (Chrysococcyx caprius) host races and their mainly ploceid hosts, suggesting that eggshell strength may also be a product of coevolution, just as egg mimicry. Still, this result depended to some extent on the inclusion of an open nesting emberizid host, which had the thinnest eggshells (Spottiswoode, 2010). Furthermore, rejection data are still unavailable for these host species preventing the crucial test of relating rejection rate to cuckoo eggshell strength. To conclude, there are many opportunities to conduct intraspecific and interspecific studies of eggshell strength and its consequences for both parasites and their hosts. Here we have emphasized the diversity of hypotheses and the degree of exclusivity of predictions. We will only be able to make progress by considering the diversity of model systems and the diversity of behavior in a coevolutionary context.
Acknowledgments
We thank two anonymous reviewers for the helpful comments on our manuscript. This work was supported by National Natural Science Foundation of China (Nos. 31071938 to AA, BGS and WL, 31272328 to WL, 31101646 and 31260514 to CY), and Program for New Century Excellent Talents in University (NCET-10-0111 to WL). BGS and FF were funded by the Research Council of Norway (218144).
Abraham S. 1973. The Kanjirankulam breeding bird sanctuary in the Ramnad district of Tamil Nadu. J Bombay Nat Hist Soc, 70: 549–552.
Ali S, Ripley SD. 1978. Handbook of the Birds of India and Pakistan. 2nd edition. Oxford University Press, Bombay.
Baker ECS. 1929. Birds: The fauna of British India including Ceylon and Burma. 2nd edition. Taylor and Francis, London.
BirdLife International. 2001. Threatened Birds of Asia: The BirdLife International Red Data Book. Part A. Cambridge, UK, BirdLife International.
BirdLife International. 2011. Species factsheet: Pelecanus philippensis. . Accessed 11 February 2011.
Brown LH, Urban KK. 1969. Breeding biology of the Great White Pelican (Pelecanus onocrotalus roseus) at Lake Shala, Ethiopia. Ibis, 111: 199–237.
Campbell WH. 1902. Nesting of the Grey Pelican in the Cuddapah district of Madras Presidency. J Bombay Nat Hist Soc, 14: 401.
del Hoyo J, Elliot A, Sargatal J. 1992. Handbook of the Birds of the World. Vol. I. Lynx Editions, Barcelona.
Ganguli U. 1964. Pelicans breeding at Mootadaippu. Newsl Birdwatcher, 4: 1–2.
Gee EP. 1960. The breeding of the Grey or Spotted billed Pelican, Pelecanus philippensis Gmelin. J Bombay Nat Hist Soc, 57: 245–251.
Islam MZ, Rahmani AR. 2004. Important Bird Areas in India: Priority Sites for Conservation. Indian Bird Conservation Network: Bombay Natural History Society, Bombay, India and BirdLife International, Cambridge, UK.
Johnson MJ. 1971. The heronry at Koonthakulam, Tirunelveli district, Tamil Nadu. Newsl Birdwatcher, 11: 1–3.
Kannan V. 2007. Nelapattu Bird Sanctuary: An Overview. Zoos Print Vol. XXII, No. 9, pp. 24
Lamba BS. 1963. Nidification of some common Indian birds. No. 7. The Spottedbilled or Grey Pelican. Pavo, 1: 110–119.
Legge WV. 1880. A History of the Birds of Ceylon. 2nd edition. Dehiwala: Tisara Prakasakayo. Sri Lanka.
Nagulu V, Rao J VR. 1990. The Grey Pelican at Nelapattu. Mayura, 12: 92–96.
Nagulu V. 1983. Feeding and breeding biology of Grey Pelican at Nelapattu Bird Sanctuary in Andhra Pradesh, India. Dissertation, Osmania University, Hyderabad, India.
Narasimhulu E. 1995. Studies on the ecology of selected coastal wetlands of Andhra Pradesh: Distribution of mangroves and associated flora and fauna with special reference to conservation strategies. Dissertation. Sri Venkateswara University, Tirupati, India.
Neelakandan KK. 1949. A South Indian Pelicanry. J Bombay Nat Hist Soc, 48: 656–666.
Oates EW. 1878. Notes on the nidification of some Burmese birds. Stray Feath, 7: 40–52.
Parr J, Eames JC, Sun H, Som H, Vi La P, Seng KH. 1996. Biological and socio-economic aspects of waterbird exploitation and natural resource utilization at Prek Toal, Tonle Sap Lake, Cambodia. IUCN Species Survival Commission, IUCN, Gland. Switzerland and Cambridge, U.K.
Rao JVR. 1986. Studies of the migratory bird, the Grey Pelican (Pelecanus philippensis) to analyse and assure ecological principles spelling survival in their traditional haunts in India during winter sojourn. Final Report, MAB Project No. 20/14/82 – MB/EN. 2. 22, July 1983.
Rao KM. 1995. Conservation of birds in coastal areas of Andhra Pradesh. In: Vijayan L (ed) Avian conservation in India. Proc. of the Avian Conservation Workshop, Coimbatore, August 1993. Salim Ali Centre for Ornithology & Natural History, Coimbatore and BirdLife Asia Council, pp 67–68.
Rhenius CE. 1907. Pelicans breeding in India. J Bombay Nat Hist Soc, 17: 806–807.
Santharam V. 1993. Nelapattu — Time to sound alarm. Mayura, 10: 34–35.
Schaller G. 1964. Breeding behaviour of the White Pelican Yellow Stone. Condor, 66: 3–23.
Schreiber RW. 1977. Maintenance behaviour and communication in the Brown Pelican. Ornithological Monograph (No: 22). American Ornith Union.
Sharma PK, Raghavaiah PS. 2002. Effect of rainfall on Grey Pelican (Pelecanus philippensis) arriving and breeding at Nelapattu Bird Sanctuary, Andhra Pradesh. Indian Forest, 128: 1101–1105.
Subramanya S, Manu K. 1996. Saving the Spot-billed Pelican a successful experiment. Hornbill, 2: 2–6.
Taher H. 2007. The Spot-billed Pelican Pelecanus philippensis of Uppalpaddu (Guntur) district, Andhra Pradesh, India. Indian Bird, 3(1): 13–16.
Talukdar BK. 1994. The status of the Spot-billed Pelican in Assam, India. Orient Bird Club Bull, 30: 13–14.
Talukdar BK. 1995a. Spot-billed Pelican in Assam. Orient Bird Club Bull, 22: 46–47.
Talukdar BK. 1995b. Pelicans fail to nest as normal at Kaziranga National Park in 1994. Oryx, 29: 157.
Venkataraman C. 1996. Human disturbance — A major factor for nest desertion. Newsl Birdwatcher, 36: 33–34.
Whistler H. 1949. Popular Handbook of Indian Birds. 4th edition. Gurney and Jackson, London, pp 489–491.
Table
1.
Summary of main selective pressure increasing eggshell thickness and major predictions of the four hypotheses for the evolution of thick eggshells in brood parasites
Laying damage hypothesis
Puncture resistance hypothesis
Chick vigour hypothesis
Anti-bacterial protection hypothesis
Main selective pressure
Damage to host eggs and prevention of damage to parasite eggs
Ejection of parasite eggs made difficult for hosts
Calcium in eggs allows precocial development of skeleto-muscular system in brood parasite chick
Thicker eggshell should provide greater protection against microorganisms
Damage to host eggs
Yes
Yes
Not important for hypothesis
Not important for hypothesis
Distance from laying cuckoo to host eggs
Dropped from distance except in small parasites that utilize hosts of similar size
Not important for hypothesis
Not important for hypothesis
Not important for hypothesis
Damage to host eggs reduces competition with host chicks
Yes
Not important for hypothesis
Not important for hypothesis
Not important for hypothesis
Damage to hole- or domenesting species common
Yes
Not important for hypothesis
Not important for hypothesis
Not important for hypothesis
Shell thickness of evicting and non-evicting parasites
No difference
No difference
Relatively thinner eggs in non-evicting parasites
No difference
Shell thickness in different host races
Relatively thicker eggs in parasite eggs from host races exploiting hosts with stronger eggshells
Relatively thicker eggs in parasite eggs from host races exploiting puncture ejecter species
Relatively thicker eggs in parasite eggs from host races exploiting hosts with larger eggs
Relatively thicker eggs in parasite eggs from host races exploiting larger hosts because larger hosts have more bacteria

 DownLoad:
DownLoad:





 Email Alerts
Email Alerts RSS Feeds
RSS Feeds