Lubna ALI, Noor-un-nisa, Syed Shahid SHAUKAT, Rafia Rehana GHAZI. 2011: Sex and age classes of prey items (rats/mice) in the diet of the Barn Owl (Tyto alba) in Sindh, Pakistan. Avian Research, 2(2): 79-86. DOI: 10.5122/cbirds.2011.0012
Citation:
Lubna ALI, Noor-un-nisa, Syed Shahid SHAUKAT, Rafia Rehana GHAZI. 2011: Sex and age classes of prey items (rats/mice) in the diet of the Barn Owl (Tyto alba) in Sindh, Pakistan. Avian Research, 2(2): 79-86. DOI: 10.5122/cbirds.2011.0012
Lubna ALI, Noor-un-nisa, Syed Shahid SHAUKAT, Rafia Rehana GHAZI. 2011: Sex and age classes of prey items (rats/mice) in the diet of the Barn Owl (Tyto alba) in Sindh, Pakistan. Avian Research, 2(2): 79-86. DOI: 10.5122/cbirds.2011.0012
Citation:
Lubna ALI, Noor-un-nisa, Syed Shahid SHAUKAT, Rafia Rehana GHAZI. 2011: Sex and age classes of prey items (rats/mice) in the diet of the Barn Owl (Tyto alba) in Sindh, Pakistan. Avian Research, 2(2): 79-86. DOI: 10.5122/cbirds.2011.0012
Vertebrate Pest Control Institute, Southern-zone Agricultural Research Centre, Pakistan Agricultural Research Council, P. O. Box-8401 Karachi University Campus, Karachi-75270, Pakistan
2.
Institute of Environmental Studies, University of Karachi, Karachi-75270, Pakistan
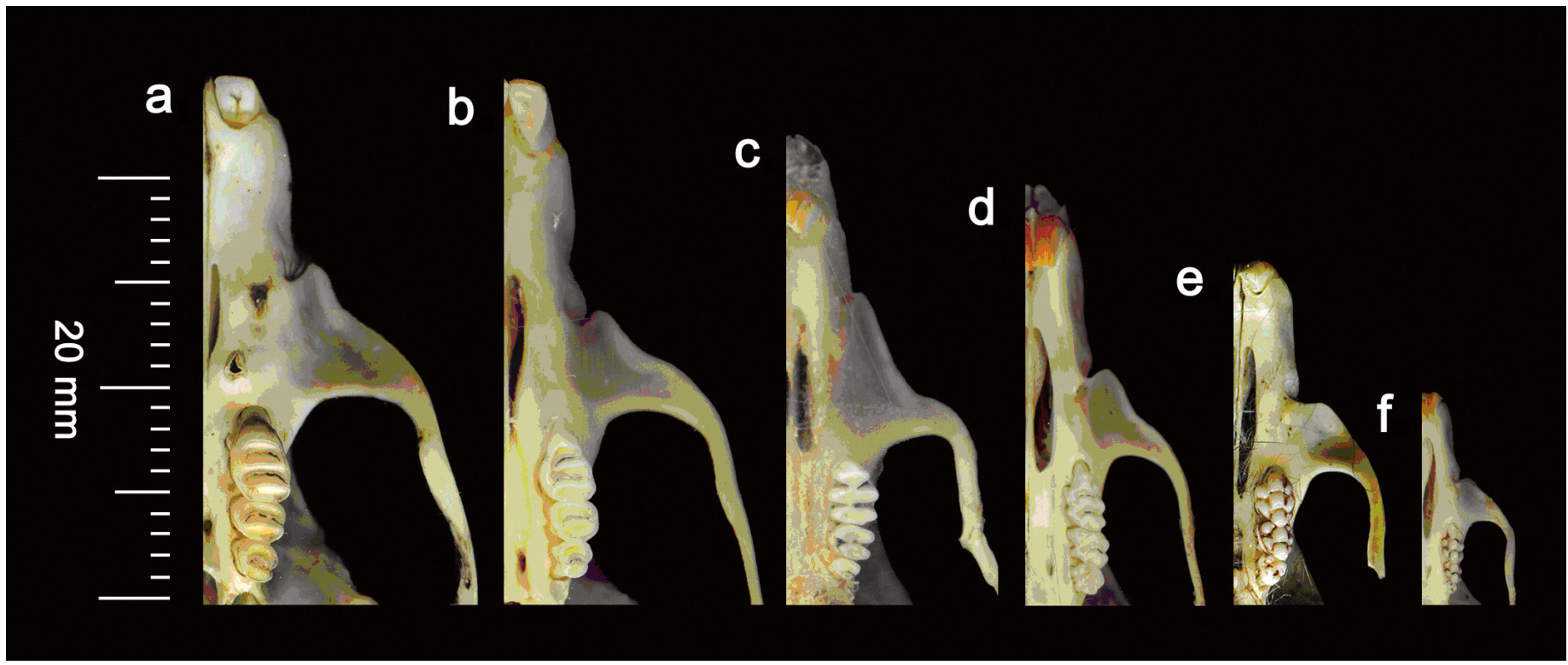
Barn Owl (Tyto alba) pellets were collected from nine locations and two districts of Sindh, Pakistan and 937 prey items were recovered from 619 pellets. Rats/mice (59.6%) were the most dominant food items consumed by the Barn Owl. Shrews (22.3%), bats (1.3%), birds (12.0%), insects (1.3%), frogs (2.2%) and plant materials (1.3%) were found in their diet as well. Study of the pelvic girdle bones of rats/mice, used only for sexing, proved to be a useful device in population dynamics. In the pelvic bone, pelvic symphysis is found only in female rats/mice developed as a result of sex hormones that occur during gestation. Among the diet of rats/mice, males were found to be significantly dominant. Tooth wear patterns on the occlusal surfaces of molariform teeth of the rats/mice were found to provide an effective criterion for establishing age classes of rats/mice. In the present study, adult rats/mice were found to be dominant over sub-adults and old adults. ANOVA showed significant differences in the number of rats/mice and shrews (prey items) and the other prey items/plant materials in the diet of Barn Owls in the district Thatta and district Karachi. Chi-square test disclosed non-significant differences in age and sex categories.
The Hooded Crane (Grus monacha) is a vulnerable species, since 2000 listed in the ICUN Red List as a threatened species. The species consists of about 11600 birds throughout the world with a declining population trend (BirdLife International, 2008). The surviving Hood Cranes breed primarily in the Amur and Ussuri river basins, along the border between Russia and China from around late July to early September (Fig. 1). Most birds head southwest towards the Three Rivers Plain in China to overwinter in Izumi in Japan (Liu et al., 2003), or along the river systems of the Songnen Plain in northeastern China to overwinter in southeastern China (Kim et al., 2010), covering more than 3000 km between their breeding and wintering areas. During their long migration, Hooded Cranes need to take one or two rest stops to replenish their expended energy en route (Bysykatova et al., 2010). The western region of the Songnen Plain receives considerable attention by Hooded Cranes for its abundant food resources and is an important net node between their breeding and overwintering sites during migration (Hiroyoshi and Johanna, 2005). Changgou village is one of the most important stopover sites in the Songnen Plain (Fig. 1), where hundreds of Hooded Cranes make a major feeding stop to refuel their fat in the spring and fall of each year.
Figure
1.
Migration routes of the Hooded Crane in the fall. The blue areas indicate stopover sites; the red circle is the location of the Changgou region; the pink areas indicate breeding sites and the end of lines with an arrow are wintering sites of the Hooded Crane.
The behavior pattern and physical condition of this migratory species might be limited by the conditions encountered at these stopover sites, except for the conditions in breeding or wintering areas (Newton, 2006). In Changgou village, most of the foraging sites for the Hooded Crane are located in arable areas and grasslands. The stopover period of the Hooded Crane is generally coincident with the cultivation or harvest season. During the plowing period or harvest season, local farmers drive their tractors/motorcycles on arable land, engaging in various agricultural activities which can cause various sound stimuli to the Hooded Crane. In addition, all year around herds of cattle are grazing in the grassland adjacent to the arable land. These cows make relatively loud moo sounds. Human stimuli, perceived as a form of risk to wildlife, is generally believed to affect the behavior pattern of wildlife in a negative sense (Frid and Dill, 2002), i.e., greater flushing distances and an increase in vigilance (Hanson and Grant, 1991) and in the long run may even affect the physical condition of the cranes. However, till now there is a lack of information on the quantitative effect of human-caused disturbances on the behavior pattern of Hooded Cranes at the stopover sites in Changgou village.
The objective of this research was to explore the effect of human disturbance on the Hooded Crane at the stopover sites in northeastern China. The results from this research will not only improve our understanding of the migration process of the Hooded Crane, but will also be useful in defining conservation strategies to sustain this vulnerable species.
Study area
The study area is located in Changgou village (47°21′43″N, 124°17′42″E) in Tiefeng District of Qiqihar City, Heilongjiang Province in northeastern China (Fig. 1). In east of Changgou area is a large wetland — Zhalong Marsh, which provides most of roosting sites for the birds. Land cover consists mainly of grassland, arable land and reed marsh (Fig. 2). Large portions of grassland and reed marsh are close to arable land but relatively far away from the local community, which makes it a favorable stopover site for migratory cranes to recover their energy after long distance travel.
Figure
2.
Foraging and roosting sites of the Hooded Crane stopover in Changgou village and the locations of four observation spots (A, B, C and D). Land structure was based on ETM+ images prevailing on 5 September, 2009.
Based on a survey on 10 September 2009, the total population in Changgou village is 2068 (Luo, 2010). Most of its residents are farmers (1230) and animal tenders (473). The remaining residents are government employees and students. The information of resident income and sources is presented in Table 1. Most sources of income originate from agricultural production and animal husbandry. The annual income of the residents is very low, at US$ 460 per capita (Table 1).
Table
1.
Gross annual income (104 US dollar) in the Changgou region
We refer to foraging sites as stopover sites at the arable land used by the birds for searching food in the daytime and roosting sites were the habitat only used for resting at night.
Methods
Sampling scheme and crane counting
Approximately 67% of the Hooded Cranes use the western part of the Songnen Plain as a stopover site, one migratory flock is at our study site and another large migratory flock of the cranes is on the eastern side of Zhalong Marsh and foraging in agricultural fields out off the wetland, following their western migratory route to the wintering sites in the south of China (Shimazaki et al., 2004; Hiroyoshi and Johnna, 2005). Generally, the Hooded Cranes arrive in Changgou village or adjacent stopping sites from their wintering site in late March and then leave in late April for their breeding sites. Each year, in mid-September, the Hooded Cranes return and leave in late October for their wintering sites. This study was conducted during the period of 2009 to 2011.
Four representative spots, referred to as A, B, C and D, were selected as observation posts for counting Hooded Cranes at the stopover sites. Detailed information of these four spots is provided in Table 2 and Fig. 2. The number of cranes was recorded simultaneously at these four spots by use of a telescope in a dark shelter in order to avoid disturbing the wildlife. Data collection was conducted from 25 March to 25 April and 20 September to 25 October every two days during 2009 to 2011. The time for observations was designed from 05:00 to 17:30 hours.
Fresh feces produced by Hooded Cranes were collected on 4 October, and 24 October in 2010 at foraging sites. The physical components of feces were then analyzed following the method by Zduniak (2005).
Auditory disturbance intensity measurements
When counting the number of Hooded Cranes, we also investigated their performance in vigilance based on the method by Treves (2000). The scanning event (number of head-up scans) and the total duration of the scans was counted in the field as vigilance events. The duration of the vigilance was measured with a stopwatch with a minimum reading of a second. The measured duration of the vigilance at these four spots before 1 October (before the harvest period) was treated as the control (non-disturbance).
Sound intensity of disturbances was measured at one meter distance from the sound source with a digital sound level meter (1350 A, Shenzhen Apparatus Technology) on 4, 8 and 15 October 2010. Sounds investigated include those made by pedestrians, voices of local farmers, noises produced by truck and tractor engines, the roar of motorcycles, mooing sounds eminated from cattle, the honking of vehicles and shouts by local people.
Hanson and Grant (1991) defined the flushing distance as the distance between a noise stimulation and the birds when they are flushed to fly away. This distance is measured as the difference in geographical location of two objects. Flushing distances of the Hooded Cranes, disturbed by sound, were also observed from 20 September to 25 October 2010.
Results and discussion
Agricultural production and foraging
Physical conditions of migratory cranes rely on encountering suitable conditions at various staging places along their routes. The migrants could suffer if conditions deteriorate at any one point. Conservation awareness, which is closely related to local standards of living and public awareness, should have a great impact on staging conditions.
The recorded dates of arrival and departure, as well as the peak date of the Hooded Crane at these four stopover sites are presented in Table 3. During recent years, hundreds of migrating Hooded Cranes have made a stop in Changgou village to replenish their body fat. On 18 October 2010, the peak of the Hooded Crane population, at 620, was recorded at their arrival in Changgou village (Table 3, Fig. 3). In 2009, staging of the Hooded Crane lasted 56 days and 63 days in 2010 (Table 3). Corn is the major food of the Hooded Crane. According to the results by Luo et al. (2010), the corn eaten by each crane is approximate 750 g per day during their staging period in Changgou village. Taking into account the total population of the Hooded Crane, the corn consumed by the birds was approximately 27 t in 2010.
Table
3.
Stopover phases of the Hooded Crane (Grus monacha) observed in 2009, 2010 and 2011
Figure
3.
Daily minimum temperatures (gray bars) in the Changgou region and the number of Hooded Cranes (empty square) stopovers in Changgou village in the fall of 2010 and in the spring of 2011. The harvest period was from early October to late October and the cultivation season from late March to early May.
Agricultural production is the major source of income for most of the local residents (Table 1). Local farmers with their low income ($460 per capita per year) are reluctant to bear the loss caused by the foraging cranes. Since local farmers are concerned about the impact of Hooded Cranes on grain production, disturbance to the cranes is inevitable during stopover periods. According to the survey by Luo et al. (2010), more than 80% of local residents will choose to drive the Hooded Crane from their arable land, should they see the cranes feeding on their crops. Hence, there is a conflict between the interest of farmers and foraging Hooded Cranes (Fig. 4). Without any economic compensation to offset the loss caused by foraging Hooded Cranes, farmers will drive this vulnerable species out of their arable land or harvest the crops as much as possible from the land, leaving almost nothing for these birds. For example, farmers started to harvest their crop earlier in order to avoid the greater loss by foraging of the peak population of Hooded Cranes arriving in late October (Table 3, Fig. 3). In 2010, farmers started to harvest the crop in early October and finished harvesting in late October. More than 60% of the crops had been harvested, when the peak population of Hooded Cranes arrived in Changgou village on 18 October 2010. The decline in food supplies at the stopover sites may lead to one of two outcomes: 1) the cranes arriving late would have to take greater risks to hunt their food from arable land; 2) cranes coming late would have to reduce their foraging rate for the rest of their migration (Newton, 2006).
Figure
4.
Food competition loop between local farmers and Hooded Cranes. The negative sign (-) represents negative feedback and the positive sign (+) represents positive feedback. Agricutural products were not only the main source of income for local farmers, but also the main food supply for the Hooded Crane. The food consumed by Hooded Cranes reduces the annual income of the farmers and affect their level of subsistence locally: the larger the number of wildlife stopover at Changgou village, the greater the amount of crops consumed by birds while, understandably local farmers want to retain as much of their crops as possible in order to improve their standard of living level; poor living condition vs. a considerable amount of agricultural products stolen by Hooded Cranes over years without any economic compensation would facilitate the conflict loop between two sides.
Human disturbance of wildlife at stopover sites has a significant effect on the feeding rates and weight gain of migrants (Drent et al., 2003). In Changgou village, the stopover period of the Hooded Cranes is essentially coincident with the cultivation or harvest season (Table 3, Fig. 3). The cultivation season is from late March to early May and the harvest period from early to late October. During the plowing period or harvest season, local farmers drive their tractors/motorcycles into arable land to engage in various agricultural activities, which can cause various sound stimuli to the Grus monacha.
Frequent human-caused disturbances may negatively affect the feeding processes of wildlife (Frid and Dill, 2002; Randler, 2006). According to field observations, most Hooded Cranes at stopover sites leave their roosting site for foraging at dawn and return to the roosting site for rest at dusk. If not disturbed, Hooded Cranes spend a large proportion of time on foraging in arable land. For example, on 24 September, the continuous whooping sound of Hooded Cranes was observed at 05:05 am, i.e., before dawn. The Hooded Cranes then left their roosting sites at 05:10 am for foraging at spot A. They returned to the roosting site at 17:30 pm when it was already dusk. Agricultural activities, especially at the harvesting period, may disturb the regular behavior pattern of Hooded Cranes. For example, on 2 October 2010, the sounds of a tractor could be heard at 04:55 am and two farmers drove a tractor roaring towards spot A at 05:30 am. The feeding process of the flock of cranes was interrupted. Whenever motorcycle(s) or tractor(s) came near a foraging site with a loud noise, we observed that the Hooded Cranes would become alarmed. Some of them would stop feeding and raise their neck scanning frequently, sometimes even hovering above the arable land for minutes. When the disturbance disappeared, they would continue to forage, otherwise, the cranes would leave the site for the next foraging site or for napping in the grassland.
Flushing distances are usually considered to be a good indicator for measuring the human-caused stimuli and for planning of buffer zones maps (Blumstein et al., 2003). The sound intensity of various auditory stimuli and flushing distances under different sound pressures are given in Fig. 5. It is expected that the louder the noise, the larger the flushing distance (Fig. 5). Foraging cranes will flush away for a distance of about 600 m when disturbed by motors, but ignore low sound noises by local people reaping on arable land at a distance of less than 300 m. Whistles and shouts in a field generated the largest sound intensity, about 120 dB, which flushed cranes over a distance above 700 m. Interestingly, the mooing sound of cattle with an intensity of 76 dB generated the shortest flushing distance. It seemed that human-caused auditory stimuli are considered a form of predation risk by the cranes, which provoke the most pronounced disturbance to Hooded Cranes.
Figure
5.
Flushing distance of the Hooded Crane disturbed by different noises. Squares are the observations (means±SE) and the red line is the Boltzmann fit. p is the noise made by pedestrians (19.6 ± 0.93; range 17–22 dB; n = 5); t is the level of voices of local people (39.7 ± 1.53; range 35–46 dB; n = 10); e is the noise from truck engines (62.5 ± 0.71; range 60–65 dB; n = 8), r is the noise from motorcycles roaring (85.4 ± 2.48; range 78 –92 dB; n = 5); h is the honking of vehicles (108.8 ± 3.71; range 98–120 dB; n = 4) and s the sound level of shouting (117.8 ± 3.47; range 110–130 dB; n = 6); y in the fitted model represents flushing distance (m) and x for the sound intensity (dB).
Comparisons of flying time, foraging time, napping time and vigilance duration between undisturbed and disturbed conditions is presented in Fig. 6. When disturbed, a large part of the daytime was spent on grassland rather than on arable land (Fig. 6a), suggesting that Hooded Cranes may have to reduce their feeding rate when disturbed. As well, frequent human stimuli alert wildlife by increasing the duration of their vigilance by more than 200% (Fig. 6b) and their flying time from 0.4 h up to 0.7 h (Fig. 6c). The longer and more frequent the vigilance, the greater the expense of sacrificing feeding during the available daytime (Fig. 6c), or indicating a signal of fear (Welp et al., 2004).
Figure
6.
Comparison of the behavior of Hooded Cranes between disturbed and undisturbed conditions. The error bar is the standard error and the sample size n is 12, 9 and 10 for (a), (b) and (c), respectively.
Frequent interruption to Hooded Cranes during the foraging period would not only exhaust the crane flock but decrease their refueling rate and might expand their stopover phase at staging areas, since the Hooded Cranes need to acquire more food than usual to finish their remaining flight and for other activities. Although competition for food may help to regulate populations in some cases, this increase in vigilance and large flushing distances of this endangered species indicate that it is imperative to define a conservation zone around sensitive areas at stopover sites.
Effect on physical condition
The physical components of feces collected on 4 October and 24 October 2010 are presented in Table 4. The crops foraged by Hooded Cranes include maize, beet tops, millet, soy bean and grass seed (Table 4). More than 90% of the feces components of Hooded Cranes consists of maize, suggesting that maize is the favorite food for these cranes at their stopover sites. Food availability at the stopover sites affects the composition of their diet. In early October, farmers start to harvest their crops, such as maize. At this time, food sources are relatively abundant. In late October, most of the crops have been harvested, while more migrating cranes stop in Changgou village to compete for a limited food supply. Shortage of maize in late October may force cranes to eat more beet tops and millet as substitutes. That is the main reason for the relative amount of maize in feces to decline from 96% on 6 October to 90% on 24 October, while the percentage of beet tops in feces significantly increased from 1.1% on 6 October to 4.5% in 24 October (p < 0.05) (Table 4).
Table
4.
Physical component and pH1:5 of crane feces collected on 4 October, 2010 and 24 October 2010
Date
Amounts
Weight (g)
Component (%)
pH1:5
Well formed
Unformed
Fresh
Air dried
Maize
Beet tops
Millet
Soy bean
Grass seed
4 October
126
5
1.8 ± 0.3
1.1 ± 0.2
96.3
1.1
0.8
2.4
0.4
7.6 ± 0.3
24 October
75
12*
1.5 ± 0.3
0.8 ± 0.2
90.6
4.5*
1.1
3.5
0.3
7.5 ± 0.4
Note: values followed by * are significantly different for the same component with paired Student t-tests (p < 0.05) between samples on 4 October and 24 October, 2010
Features of feces left by wildlife are good indicators of their physical conditions (Vilés, 2004; Zduniak, 2005). Generally, the average weights of fresh and dried feces are relatively higher when food sources are abundant in early October: in the case of our Hooded Cranes, the average weights of fresh and dried feces were 1.8 and 1.1 g on 6 October and 1.5 and 0.8 g on 24 October. There were significantly higher percentages of unformed feces on 24 October (12/87) than on 6 October (5/131), which might indicate that health conditions of the Grus monacha arriving late were inferior to those arriving early. When food becomes scarce, crane flocks will have to regulate their diet, leave for the next stopping site or fly directly towards their destination, which, in the long run, might reduce the Hooded Crane population. It was not clear whether the inferiority of the body condition of the Hooded Cranes arriving in late October was related to adverse weather conditions. On 24 October, 2010 the minimal temperature reached −9.0℃ (Fig. 3).
Conservation considerations
According to the theory of system dynamics, an isolated system can be transformed into an open system when outside matter and energy from the environment is introduced into the system. Some successful experiences in other breeding or overwintering sites may be utilized as reference to break the competition loop. For example, the Zhanhe National Natural Reserve of China not only compensates local farmers for food loss caused by the Hooded Crane, but also encourages and awards people who leave enough maize on arable lands close to breeding sites (Guo et al., 2005). In this way, the Hooded Crane can obtain a large enough food supply (Hiroyuki, 2004). In addition, defining a buffer zone around the stopover sites during the migratory phase could help reduce the disturbance by various human-caused acoustic pressures (Rodgers and Smith, 1997). On the basis of flushing distances and the location of main foraging and/or resting sites for Hooded Crane flocks, a buffer zone for a reserve scheme was mapped using the buffer tool of Arcgis software (version 9.1) (Fig. 7). According to this mapping scheme, various human-caused disturbances should be prohibited in the core zone and their level and frequency controlled in the buffer zone during the stopover period of the species until the migratory cranes had left the stopover site.
Figure
7.
Proposed core and buffer zone for the conservation of Hooded Cranes. The core area is about 20 km2 and the buffer zone 30 km2.
Education programs are urgently required at stopover sites to promote public awareness of the status of this species. Special compensation funds by local government at stopover sites for the crops consumed by Hooded Cranes are urgently required to solve the food conflict between local farmers and Hooded Cranes. With the participation of local government and farmers, the competition loop could be transformed into a sustainable development system as depicted in Fig. 8.
Figure
8.
Sustainable development mode between local farmers and the Hooded Crane. The negative sign (−) represents negative feedback and the positive sign (+) represents positive feedback. Positive practices for conservation comprise not only special funds by local governments for the crop consumed by Hooded Cranes but also rewarding those people who leave enough agricultural products on arable land for this species of birds. Employment practice i.e., guiding surplus-labor work in urban areas would not only improve the standard of living, but decrease human stimuli to the migrant cranes as well. Positive practices and repositioning of personnel policy would bring about more wealth locally and a more suitable stopping habitat for wildlife as well. Given such conditions, more migrant cranes would choose to stopover in the Changgou region, unless constrained by the capacity of its environment.
Our results indicate that the conflict between the interest of farmers and foraging Hooded Cranes not only brings about various human disturbance to this vulnerable species, but also limit the food supply available for the migratory cranes, affecting their health, especially of cranes arriving late. Frequent agricultural activities in the field produce strong disturbances to foraging Hooded Cranes. Human-caused disturbances not only frequently interrupt the feeding process of the cranes, but also lead to a 200% increase in vigilance and an increase in flying time from 0.4 to 0.7 h. High-intensity sound pressure will cause larger flushing distances. Whistles and shouts in the field generated the largest sound intensity, about 120 dB, which disturbed the cranes at distances of over 700 m.
Food availability at the stopover sites affected the composition of the diet, as well as the physical condition of this species of wildlife. In late October, most of the crops have been harvested and more migrating cranes stop in Changgou village to compete for limited food. A shortage of maize in late October may force cranes to eat more beet tops and millet as substitutes, thereby affecting their health, as indicated by a significantly higher percentage of unformed feces and, to some extent, as the result from a change in diet.
To reduce the effect of human-caused disturbances on the Hooded Cranes, it is imperative that a buffer zone be defined around sensitive areas during stopover periods. In addition, to solve the conflict between the interest of farmers and the demand for crops by the Hooded Crane, financial compensation to the farmers for the crops consumed by Hooded Cranes is imperative for a better protection of this vulnerable birdlife.
Acknowledgements
We would like to extend our appreciation to the undergraduate students in the Department of Geography Science at Qiqihar University for their field work.
Alivizatos H, Goutner V, Zogaris S. 2005. Contribution to the study of four owl species (Aves, Strigiformes) from mainland and island areas of Greece. Belg J Zool, 135: 109–118.
Alvarez-Castaňda ST, Carenas N, Mendez L. 2004. Analysis of mammal remains from owl pellets (Tyto alba), in a suburban area in Baja California. J Arid Environ, 59: 59–69.
Brown JS, Kotler BP, Smith RJ, Wirtz, WO. 1988. The effects of owl predation on the foraging behavior of heteromyid rodents. Oecologia, 76: 408–415.
Campbell RW, Manuwal DA, Harestad AS. 1987. Food habits of the Common Barn Owl in British Columbia. Can J Zool, 65: 578–586.
Clarke JA. 1983. Moonlight's influence on predator/prey interaction between short-eared owls (Asio filammeus) and deermice Peromyscus maniculatus. Behav Ecol Sociobiol, 13: 205–209.
Dickman CR, Daly SEJ, Connell GW. 1991. Dietary Relationships of the Barn Owl and Australian Kestrel on Islands off the Coast of Western Australia. Emu, 91(2): 69–72.
Dunmire WW. 1955. Sex dimorphism in the pelvis of rodents. J Mammal, 36(3): 356–361.
Escarlate-Tavares F, Pessoa LM. 2005. Bats (Chiroptera, Mammalia) in Barn Owl (Tyto alba) pellets in northern pantanal, Mato Grosso, Brazil. Mastozool Neotrop Mendoza, 12(1): 61–67.
Glue DE. 1971. Avian predator pellet analysis and the mammalogist. Mammal Rev, 21: 200–210.
Goutner V, Alivizatos H. 2003. Diet of the barn owl (Tyto alba) and little owl (Athene noctua) in wetlands of northeastern Greece. Belg J Zool, 113: 15–22.
Greene EC. 1935. Anatomy of the Rat. Manufactured by the Haddan Craftsmen, INC. CamDen, New Jersey, U.S.A.
Gwynne DT. 1987. Sex-biased predation and the risky mate-locating behaviour of the tick-tock cicadas (Homoptera: Cicadidae). Anim Behav, 35: 571–576.
Hall K, Newton WH. 1946. The normal course of separation of the pubes in pregnant mice. J Physiol, 104: 346–352.
Hall K. 1947. The effects of pregnancy and relaxin on the histology of the pubic symphysis in the mouse. J Endocrinol, 5: 174–182.
Huebschman JJ, Freeman PW, Genoways HH, Gubanyi JA. 2000. Observations on small mammals recovered from owl pellets from Nebraska. Prairie Nat, 32(4): 209–215.
Ingels C. 1995. Summary of California studies analyzing the diet of barn owls. Sustain Agr, 7(2): 14–16.
Iqbal HM. 1975. Evaluation of the degree of tooth wear and morphometric changes as a criterion of determining age groups in soft-furred field rat (Millardia meltada). M. Sc. Thesis. Department of Zoology, University of Agriculture, Lyallpur, Pakistan, pp 60.
Jorgensen EE, Sell SM, Demarais S. 1998. Barn owl prey use in Chihuahuan Desert Foothills. Southwest Nat, 43(1): 53–56.
Khan HA, Beg MA, Khan AA. 1994. Food of Barn Owl (Tyto alba) in Lower Sindh. Pak J Agr Sci, 31(3): 233–235.
Kuno E. 1987. Principles of predator-prey interaction in the oretical, experimental and natural population systems. Adv Ecol Res, 66: 1426–1438.
Lekunze LM, Ezealor AU, Aken'Ova T. 2001. Prey groups in the pellets of the barn owl Tyto alba (Scopoli) inthe Nigerian savanna. Afr J Ecol, 39: 38–44.
Longland WS, Jenkins SH. 1987. Sex and age affect vulnerability of desert rodents to owl predation. J Mammal, 68: 746–754.
Marti CD, Hogue JG. 1979. Selection of prey size in Screech Owls. Auk, 96: 319–327.
Morris P. 1979. Rats in the diet of the barn owl (Tyto alba). J Zool Proc Zool Soc Lond, 189: 540–545.
Morse DH. 1980. Behavioral Mechanisms in Ecology. Harvard University Press, Cambridge, MA, U.S.A.
Mushtaq-ul-Hassan M, Ghazi RR, Noor-un-Nisa. 2007. Food preference of the Short-Eared Owl (Asio flammeus) and Barn Owl (Tyto alba) at Usta Muhammad, Balochistan, Pakistan. Turk J Zool, 31: 91–94.
Mushtaq-ul-Hassan M, Mahmood-ul-Hassan M, Beg MA, Khan AA. 1999. Reproduction and abundance of the house shrew suncus murinus in the villages and houses of central Punjab (Pakistan). Pak J Zool, 29: 199–202.
Mushtaq-ul-Hassan M, Raza MN, Shahazadi B, Ali A. 2004. The diet of barn owl (Tyto alba) from canal bank, canal rest house and graveyard of Gojra. J Res Sci, 15(3): 291–296.
Roberts TJ. 1997. The Mammals of Pakistan. Vol. 1. Oxford University Press, Oxford, U.K.
Roberts TJ. 2005. Field guide to the Small Mammals of Pakistan. Oxford University Press, Oxford. pp 280.
Seçkin S, Coşkun Y. 2005. Small mammals in the diet of the Long-eared Owl, Asio otus, from Diyarbakýr, Turkey. Zool Mid East, 35: 102–103.
Seçkin S, Coşkun Y. 2006. Mammalian Remains in the Pellets of Long-eared Owls (Asio otus) in Diyarbakir Province. Turkey. Turk J Zool, 30: 271–278.
Shehab AH, Al Charabi SM. 2006. Food of the Barn Owl Tyto alba in the Yahmool Area, Northern Syria. Turk J Zool, 30: 175–179.
Sih A, Crowley P, McPeek M, Petranka J, Strohmeier K. 1985. Predation, competition, and prey communities: a review of field experiments. Ann Rev Ecol Syst, 16: 269–311.
Smith DG, Wilson CR, Frost HH. 1972. Seasonal habits of barn owls in Utah. Great Basin Nat, 32: 229–234.
Tores M, Yom-Tov Y. 2003. The Diet of the Barn Owl (Tyto alba) in the Negev Desert. Isr J Zool, 49: 233–236.
Tuttle MD, Ryan MJ. 1981. Bat predation and the evolution of frog vocalizations in the neotropics. Science, 214: 677–678.
Twente JW, Baker RH. 1951. New records of mammals from barn owl pellets. J Mammal, 32: 120–121.
Yom-Tov Y, Wool D. 1997. Do the contents of barn owl pellets accurately represent the proportion of prey species in the field? Condor, 99: 972–976.
Zaman R. 1975. Age determination, age structure, sex ratio and age specific reproduction in Tatera indica (Hardwicke, 1807) population in the Punjab. Thesis, University of Agriculture, Faisalabad, Pakistan.
Zar JH. 1996. Biostatistical Analysis. 3rd edition. Prentice Hall, Upper Saddle River, New Jersey, USA, pp 662.
Table
4.
Physical component and pH1:5 of crane feces collected on 4 October, 2010 and 24 October 2010
Date
Amounts
Weight (g)
Component (%)
pH1:5
Well formed
Unformed
Fresh
Air dried
Maize
Beet tops
Millet
Soy bean
Grass seed
4 October
126
5
1.8 ± 0.3
1.1 ± 0.2
96.3
1.1
0.8
2.4
0.4
7.6 ± 0.3
24 October
75
12*
1.5 ± 0.3
0.8 ± 0.2
90.6
4.5*
1.1
3.5
0.3
7.5 ± 0.4
Note: values followed by * are significantly different for the same component with paired Student t-tests (p < 0.05) between samples on 4 October and 24 October, 2010


 DownLoad:
DownLoad:












 Email Alerts
Email Alerts RSS Feeds
RSS Feeds