Fasheng ZOU, Guizhu CHEN, Qiongfang YANG. 2010: Impacts of bird abundance, activity height and light intensity on the number of birds captured by mist netting. Avian Research, 1(4): 221-229. DOI: 10.5122/cbirds.2010.0017
Citation:
Fasheng ZOU, Guizhu CHEN, Qiongfang YANG. 2010: Impacts of bird abundance, activity height and light intensity on the number of birds captured by mist netting. Avian Research, 1(4): 221-229. DOI: 10.5122/cbirds.2010.0017
Fasheng ZOU, Guizhu CHEN, Qiongfang YANG. 2010: Impacts of bird abundance, activity height and light intensity on the number of birds captured by mist netting. Avian Research, 1(4): 221-229. DOI: 10.5122/cbirds.2010.0017
Citation:
Fasheng ZOU, Guizhu CHEN, Qiongfang YANG. 2010: Impacts of bird abundance, activity height and light intensity on the number of birds captured by mist netting. Avian Research, 1(4): 221-229. DOI: 10.5122/cbirds.2010.0017
To understand better the impact on mist net capture rates of the activity height of birds, light intensity, bird richness and abundance, we compared data from mist nets and point counts at the Jianfengling Forest Area, Hainan Island. Mist-nets were operated for a total of 7135 net-hours. A total of 587 individuals of 45 bird species were captured. The total mean capture rate was 8.6 ±10.4 per 100 net-hours. A total of 4932 individuals of 107 bird species were recorded by visual observation alone and 7616 individuals of 120 species were recorded in a visual and aural combination. The Grey-cheeked Fulvetta (Alcippe morrisonia) was the most abundant species, accounting for 38.7% of total captures, 27.9% of visual observations and 22.2% of visual + aural observations. The capture number was correlated with the number observed. Thirty-one species were more likely to be captured than observed. Canopy species, such as members of the Corvidae and Dicruridae, were seldom captured. For all ground feeding species, the capture number was greater than that of observation. Some cryptic species, such as the Lesser Wren Babbler (Napothera epilepidota), Spotnecked Babbler (Stachyris striolata) and Cuckoo Owl (Glaucidium cuculoides) also had a higher relative capture rate than that of observations. There was a significant relationship between light intensity and the number of birds captured.
Hainan Island is off the coast of southern China and home to a growing list of endemic avian species. Currently the biota of the island is under considerable threat from habitat destruction, providing a need for accurate surveys of its avifauna. The present study examined the relative effectiveness of mist netting in characterizing the forest avifauna of Hainan through the following metrics: 1) the relationship between visual and aural observations based on point counts and captures from mist netting in avian richness and abundance, 2) the relationship between activity height and capture rate and 3) the relationship between light intensity and capture rate.
Methods
Study area
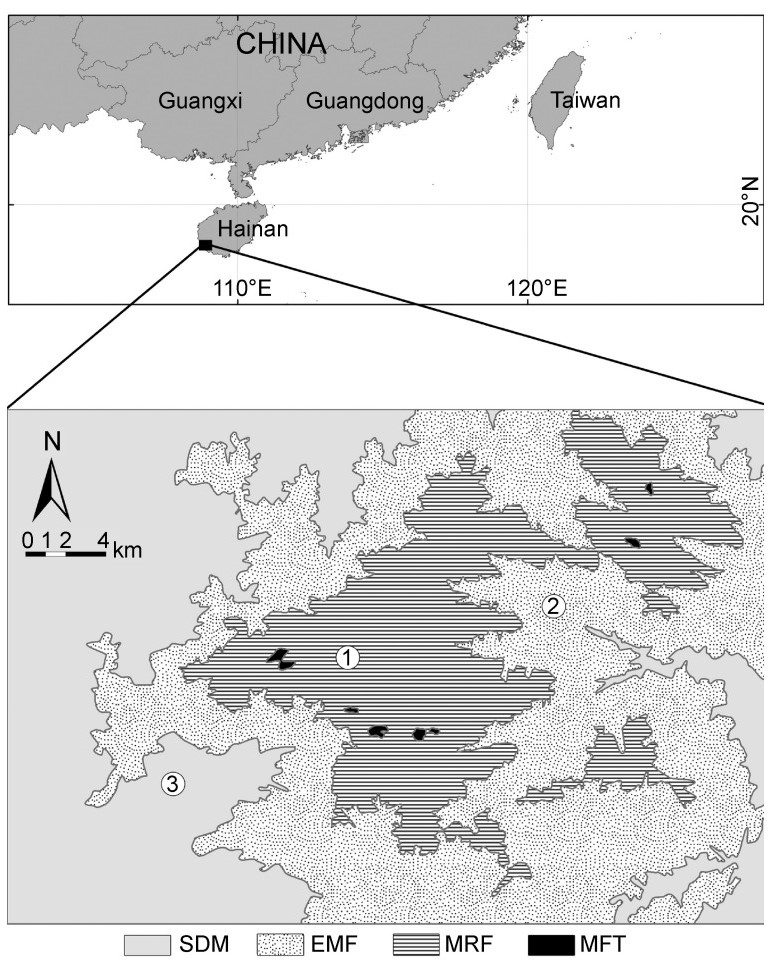
The study was conducted at the Jianfengling Forest Area (18°23′–18°52′N, 108°36′–109°05′E), which covers 600 km2 in southwestern Hainan Island and ranges in elevation from sea level to 1413 m (Fig. 1). The climate is that of a tropical monsoon area, with a wet season from May to October and a dry season from November to April. The mean annual temperature is 24.5℃ and the annual rainfall 1600–2600 mm.
Figure
1.
Sample sites of three vegetation types for semi-deciduous monsoon forest (SDM), evergreen monsoon forest (EMF) and montane rain forest (MRF). Mossy forest on top of the mountain (MFT) was not sampled (see text). Numbers 1, 2 and 3 represent sampling locations in MRF, EMF and SDM, respectively.
Jianfengling Forest Area has more than 2800 plant species and four vegetation types (Fig. 1): 1) a semi-deciduous monsoon forest (SDM) in the foothills and mountains at elevations between 100 and 400 m, 2) an evergreen monsoon forest (EMF) primarily in a hillside belt at 300–700 m elevation with a high abundance of trees of the genus Vatica, a predominant component of tropical rainforests in Asia, 3) a tropical montane rainforest (MRF) with the highest tree species richness of the four vegetation types and occurs mainly at 600–1100 m elevation, including a large number of old and tall trees with large plank buttresses and a great variety of epiphytes, parasitic vines and creepers, and 4) a mossy forest (MFT) covering a very small area on foggy mountain tops and ridges over 1100 m (Zeng et al., 1997).
Bird sampling
Birds were sampled on eight occasions, four during the wet season (May 2000, June 2001, October 2001 and September 2004) and four during the dry season (November 2000, February 2001, January 2002 and March 2002) and in three vegetation types (MRF, EMF and SDM). Birds of the mossy forest were not sampled because of their small size and access is difficult. Bird surveys in each vegetation type were conducted using fixed-radius (30 m) point counts and mist nets. At least 20 points were established on three preexisting trails for each type. All point count locations were identified by GPS (Global Positioning System, Garmin corporation, accuracy < 10 m) and the mean linear distance between nearest points in the same trail was 184.0 ± 6.5 m (n = 54). During point count periods, each point was surveyed for 10 min between 06:30 and 10:30 hours. All birds detected visually or aurally within approximately 30 m of the observer were recorded along with the estimated distance between the birds and the observer and the activity height of the bird. All points of each type were surveyed within a three-day period provided there was no strong wind or rain. Mist nets of 12 m in length, 2.6 m in height and with 36 mm mesh were used to capture birds. The center of each net site was recorded using GPS (Table 1). Nets were installed where vegetation and topography permitted and all nets were in the forest interior. Ten nets were operated simultaneously at each site. The same sampling trails and net sites were used throughout the study period. Nets were opened for 3–4 consecutive days between approximately 06:30 and 17:30 hours on days without rain or strong wind for each period. Nets were checked at intervals of about one hour. Birds were banded using numbered aluminum leg-rings, freely supplied by the National Bird Banding Center of China. After banding, all birds were immediately released at the capture site to minimize disruption of their normal movements. Time, location, species and ring code (for recaptured birds) were recorded. Light intensity was measured by DER EE Digital Light Meter (Liquid Crystal Display, made in Shanghai) at 08:00, 10:00, 12:00, 14:00 and 16:00. Five points (0, 3, 6, 9 and 12 m along the net) were selected for each net.
The migratory status of birds was based on Xu et al. (1983). "Abundant species" were those representing more than 10% of total captures (Zheng, 1995). "Rare species" were those representing less than 2% of total captures (Karr, 1971). All data were examined for normality using Kolmogorov-Smirnov tests. The relationships between observation numbers and capture numbers and light intensity and capture numbers were determined by Spearman's rank correlation coefficient. The nonparametric Kruskal-Wallis test was used to test differences of light intensity between net-days. All analyses were conducted with SPSS 16.0 except for the rarefaction analysis. EcoSim was used to compute species accumulation curves based on rarefaction (Gotelli and Entsminger, 2001).
Results
Mist netting data
Mist nets were operated for a total of 7135 net-hours. A total of 587 individuals representing 45 bird species were captured. A species accumulation curve predicted very slow growth for further sampling. The total mean capture rate during eight survey periods was 8.6 ± 10.4 per 100 net-hours (n = 237). Passerines dominated the understory avian community, accounting for 86.7% of all species and 96.9% of total captures. The only species accounting for more than 10% of total captures was the Grey-cheeked Fulvetta (Alcippe morrisonia), which accounted for 38.7% of total captures. Rare species comprised 75.6% of species. Resident birds were the major component of the understory bird community, accounting for more than 80.0% of all species and 93.7% of total captures. Only nine long-distance migratory species (including one passage migrant and eight winter visitors) were captured (Table 2). Migratory birds accounted for a small proportion of total captures (6.3%).
Table
2.
Species list of birds captured at Jianfengling a
Common name
Scientific name
Captures b
NV c
NT d
M e
Activity height (m) f
Emerald Dove
Chalcophaps indica
4
−
−
R
−
Cuckoo Owl
Glaucidium cuculoides
8
4
20
R
10.7 ± 3.0 (3)
Collared Pygmy Owl
Glaucidium brodiei
1
4
17
R
11.7 ± 3.3 (3)
Red-headed Trogon
Harpactes erythrocephalus
2
25
45
R
9.9 ± 1.2 (17)
Blue-bearded Bee-eater
Nyctyornis athertoni
1
16
47
R
14.9 ± 4.9 (7)
Black-browed Barbet
Megalaima oorti
2
13
158
R
13.5 ± 2.4 (10)
Silver-breasted Broadbill
Serilophus lunatus
26
66
68
R
12.0 ± 1.4 (12)
Blue-rumped Pitta
Pitta soror
3
3
3
R
0 (2)
White-throated Bulbul
Alophoixus pallidus
46
265
480
R
8.3 ± 1.9 (105)
Green-winged Bulbul
Hypsipetes mcclellandii
5
28
52
R
12.2 ± 1. 8 (13)
Chestnut Bulbul
Hemixos castanonotus
5
123
354
R
11.3 ± 3.0 (27)
Black Drongo
Dicrurus macrocercus
1
15
19
R
16.0 ± 1.0 (8)
Bronzed Drongo
Dicrurus aeneus
5
95
139
R
16.6 ± 4.2 (41)
Greater Racket-tailed Drongo
Dicrurus paradiseus
2
37
43
R
12.6 ± 2.1 (15)
Yellow-breasted Green Magpie
Cissa hypoleuca
2
−
−
R
−
Gray Treepie
Dendrocitta formosae
1
9
30
R
13.0 (5)
Racket-tailed Treepie
Temnurus temnurus
1
12
13
R
14.9 ± 2.4 (7)
Red-tailed Robin
Luscinia sibilans
8
1
1
W
0.5 (1)
Red-flanked Bush Robin
Luscinia cyanurus
9
2
2
W
5.0 (1)
White-rumped Shama
Copsychus malabaricus
11
16
19
R
3.9 ± 1.3 (6)
White-crowned Forktail
Enicurus leschenaulti
13
30
86
R
0 (22)
Orange-headed Ground Thrush
Zoothera citrina
11
−
−
R
−
Golden Mountain Thrush
Zoothera dauma
2
1
1
W
0 (1)
Grey Thrush
Turdus cardis
12
4
4
W
0 (1)
Blue-and-white Flycatcher
Cyanoptila cyanomelaena
1
1
1
W
−
Fujian Niltava
Niltava davidi
1
1
1
W
−
Hainan Blue Flycatcher
Cyornis hainanus
13
37
81
R
4.0 ± 0.5 (26)
White-throated Fantail
Rhipidura albicollis
4
19
24
R
4.5 ± 0.8 (14)
Black-naped Monarch
Hypothymis azurea
13
32
33
R
8.8 ± 1.3 (21)
Lesser Necklaced Laughingthrush
Garrulax monileger
4
5
6
R
6.5 ± 5.0 (2)
Greater Necklaced Laughingthrush
Garrulax pectoralis
3
9
22
R
8.1 ± 2.1 (5)
Grey Laughingthrush
Garrulax maesi
1
23
129
R
3.7 ± 1.7 (3)
Black-throated Laughingthrush
Garrulax chinensis
8
42
197
R
3.0 ± 0.9 (15)
Large Scimitar Babbler
Pomatorhinus hypoleucos
1
2
9
R
0.3 (1)
Rufous-necked Scimitar Babbler
Pomatorhinus ruficollis
42
46
217
R
1.1 ± 0.3 (18)
Lesser Wren Babbler
Napothera epilepidota
3
2
2
R
1.2 (1)
Rufous-capped Babbler
Stachyris ruficeps
17
79
142
R
4.9 ± 1.2 (19)
Spot-necked Babbler
Stachyris striolata
7
2
2
R
0.5 (1)
Gould's Fulvetta
Alcippe brunnea
23
73
258
R
2.1 ± 0.4 (28)
Grey-cheeked Fulvetta
Alcippe morrisonia
227
1376
1687
R
3.5 ± 0.2 (178)
White-bellied Yuhina
Erpornis zantholeuca
24
459
553
R
7.5 ± 0.6 (74)
Grey-headed Parrotbill
Paradoxornis gularis
10
112
112
R
10.3 ± 2.5 (6)
Dusky Warbler
Phylloscopus fuscatus
1
2
2
W
6.0 (1)
Pale-legged Leaf Warbler
Phylloscopus tenellipes
2
3
3
P
3.5 ± 1.5 (2)
Blyth's Leaf Warbler
Phylloscopus reguloides
1
−
−
W
−
a Classification based on Zheng (2005). b Captures: capture number. c NV: Visual observation number. d NT: Total observation number (visual + aural). e Migratory status based on Xu et al. (1983): resident (R), winter visitor (W) and passage migrant (P). f Sample number in brackets.
Detections by point counts and consistency with mist netting data
During point counts, a total of 107 bird species comprising 4932 individuals were visually recorded and 120 species comprising 7616 individuals were recorded visually and aurally. The Grey-cheeked Fulvetta was the most dominant species, accounting for 27.9% of visual observations and 22.2% of total observations (visual and aural detections). Significant relationships were found between the number of visual observations and the number of captures (rs = 0.61, n = 41, p < 0.01), between the number of visual + aural observations and the number of captures (rs = 0.53, n = 41, p < 0.01) and between the number of visual observations and the number of visual + aural observations (rs = 0.94, n = 41, p < 0.01). For 31 species, the relative captures were greater than the relative observations. For the Collared Pygmy Owl (Glaucidium brodiei), the Black-browed Barbet (Megalaima oorti) and the Black-throated Laughingthrush (Garrulax chinensis), the relative capture number was higher than that of visual observation during point counts, but lower than the visual and aural observations combined, indicating that these species are more readily heard than they are either seen or caught by mist nets.
Relationship between activity height and number of captures
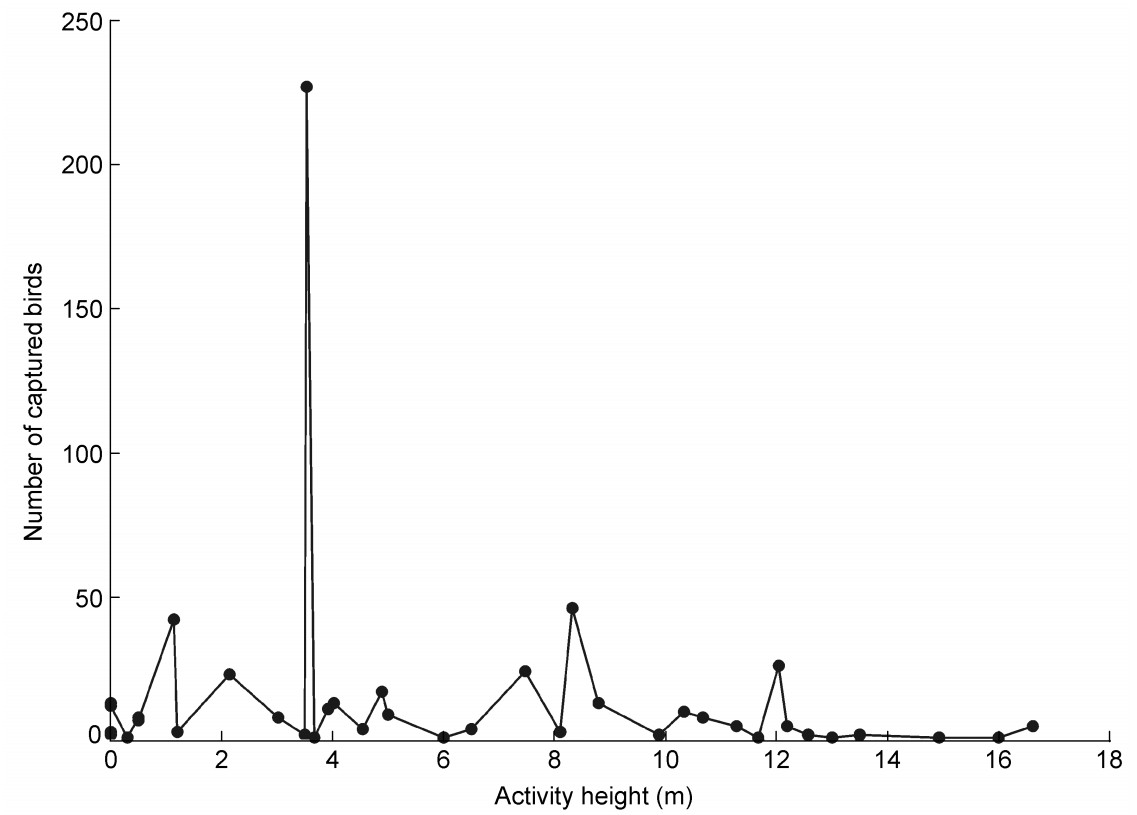
Based on point counts for 107 species, activity heights ranged from the ground, for species such as the Blue-rumped Pitta (Pitta soror), the Orange-headed Ground Thrush (Zoothera citrina) and the Golden Mountain Thrush (Zoothera dauma) to 35 m, for species such as the Chestnut Bulbul (Hemixos castanonotus), the Black Bulbul (Hypsipetes leucocephalus) and the Ratchet-tailed Treepie (Temnurus temnurus). Of the 45 species captured, 39 were also detected by point counts, where the activity height of these 39 species ranged from the ground to 17 m, averaging 7.2 ± 0.3 m (n = 697). Most species netted were mainly active below 4 m in height, but in addition there were two small peaks of captured numbers at 8.3 and 12.0 m (Fig. 2). These were mainly White-throated Bulbuls (Alophoixuspallidus, most active at 8–9 m) and Silver-breasted Broadbillds (Serilophus lunatus, 11–13 m). Many individuals of these two species were captured.
Figure
2.
Relationship between activity height and number of birds captured for 39 species recorded for both observations and mist-netting
Relationship between light intensity and number of captures
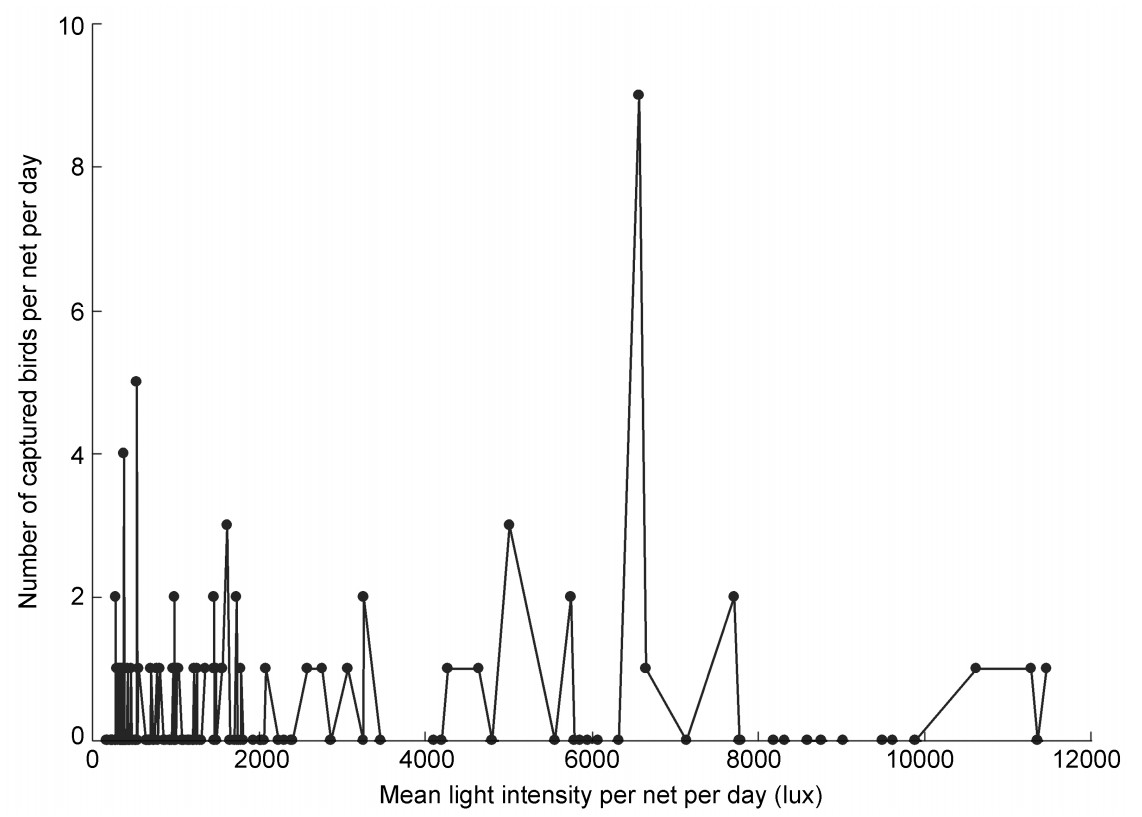
Light intensities were measured for 180 net-days during October 2001, January and March 2002, and September 2004. Light averaged 2040.6 ± 55.5 lux (n = 4500) and ranged from 1.7 to 19970.0 lux. The number of birds captured for the 180 net-days was significantly correlated with light levels (rs = 0.16, n = 180, p = 0.03) (Fig. 3). The correlation was still significant after deleting the highest point (light intensity: 6564.4 lux, captures: 9 individuals) in Fig. 3 (rs = 0.15, n = 179, p = 0.05).
Figure
3.
Relationship between mean light intensity and number of birds captured for 180 net-day measures (basic unit "net-day" = per net per day).
Critical to understanding avian abundance and diversity is the availability of comparatively unbiased tools for assessing avian communities. This is particularly important on the island of Hainan where the avifauna is poorly known and under considerable threat from anthropogenic disturbance. One test of the efficacy of a particular survey method is to measure its results against an independent alternative methodology. Congruent results from two or more independent methodologies can indicate that a particular survey method is comparatively unbiased relative to other methods.
Our study at the Jianfengling Forest Area showed a relationship between the number of observations during point counts and the number of mist net captures. Certain species (e.g. the Grey-cheeked Fulvetta) were abundant in both point count surveys and mist net captures, indicating that both techniques provided similar data regarding identification of the most abundant species. The White-throated Bulbul, Rufous-necked Scimitar Babbler (Pomatorhinus ruficollis), Silver-breasted Broadbill, White-bellied Yuhina (Erpornis zantholeuca), Gould's Fulvetta (Alcippe brunnea) and Rufous-capped Babbler (Stachyris ruficeps) were also identified as common species in both mist net and point count surveys. The Dusky Warbler (Phylloscopus fuscatus), Large Scimitar Babbler (Pomatorhinus hypoleucos) and Blue-and-white Flycatcher (Cyanoptila cyanomelaena) were identified as uncommon in both mist net and point count surveys. These data strongly support the conclusion that point counts and mist netting are providing similar measures and comparable data of avian abundance and diversity for some migratory bird species (Wang and Finch, 2002).
However, there were some important differences between mist netting and point count data at Jianfengling. Resmen and Good (1996) pointed out that activity height strongly affects the probability of capture. Our results support this conclusion. The probability of capture was greater than that of observation for ground feeders, among which, the White-crowned Forktail (Enicurus leschenaulti) and Grey Thrush (Turdus cardis) had high capture ratios. In contrast, canopy dwelling corvid and dicrudid species were seldom captured. Other canopy and aerial feeding species were more likely to be observed than captured, examples being the Blue-bearded Bee-eater (Nyctyornis athertoni) and Black-browed Barbet. These results are consistent with results from other avian communities showing that aerial or canopy-foraging species are often missed by mist netting (Ralph et al., 1993; Rappole et al., 1998; Wang and Finch, 2002). However, while mist netting underperforms relative to point counts for canopy species, mist netting is likely more effective than point counts in detecting mid-story species. Whitman et al. (1997) found that in northern Belize mist nets detected a greater proportion of mid-story species than point counts.
The effect of ambient light on the probability of capture has not been studied. Normally, strong light would make it much easier for birds to see the nets and hence avoid capture, which is the rationale behind the practice of early morning (i.e. low-light) mist netting for open country, grassland and shorebirds species. Our data showed a positive correlation between capture number and light intensity. One possibility is that areas of comparatively higher light are typical areas of disturbance and forest edge habitat. Edge specialists are particularly active in these areas since their movements are confined to narrow corridors making them more likely to run into mist nets. In contrast, at low light levels, interior forest species whose activity is less confined to narrow corridors would be less likely to encounter mist nets. Another possibility is that strong light helps birds see their food (e.g. insects) more clearly in forest and that the frequency of catching insects is greater in strong light forests than in dense, dark forests.
For cryptic, nocturnal or crepuscular species or those which infrequently vocalize, point counts are likely to underestimate abundance. Cryptic species such as the Lesser Wren Babbler (Napothera epilepidota), Spot-necked Babbler (Stachyris striolata) and the nocturnal Cuckoo Owl (Glaucidium cuculoides) were more likely to be captured than detected during point counts. Five other species, known from only a very small number of records prior to this survey (Cheng and Tan, 1973; Zeng et al., 1995), were detected by mist netting, i.e., the Red-tailed Robin (Luscinia sibilans), Golden Mountain Thrush, Lesser Wren Babbler, Pale-legged Leaf Warbler (Phylloscopus tenellipes) and Fukien Niltava (Niltava davidi). These results and those from other studies support the conclusion that mist netting is a particularly appropriate technique for sampling species that are difficult to see and seldom vocalize (Terborgh et al., 1990; Wang and Finch, 2002).
Detection by mist nets is affected by a large number of factors, such as habitat heterogeneity, flight distance and frequency (Remsen and Good, 1996). Canopy species are likely to be captured more often in nets among short forest trees than among tall forest trees, even though the species is equally abundant in both habitats (Petit et al., 1992). In other studies, canopy or sub-canopy species were detected by mist net more often in the young forest habitats than in the mature forest habitat (Pagen et al., 2002). Mesh size may also impact capture rate. Piratelli (2003) concluded that 36 mm mesh nets were better than 61 mm mesh nets for most birds. However, for our study site at Jianfengling, mist netting and point counts provided largely congruent results with a few notable exceptions. Robust methods for estimating avian diversity and abundance are especially important in avian communities of subtropical and tropical Asia, as on Hainan Island, where data on threatened populations are sorely needed.
Acknowledgements
We are grateful to the Department of Wildlife Conservation and Management, the Hainan Forest Bureau and the Jianfengling Forest Bureau for permission to conduct fieldwork at the Jianfengling Forest Area. We thank Yayun Wang, Nin Guo and Huanqiang Chen for help during fieldwork. We thank James Van Remsen and Herman Mays for providing helpful comments which improved the manuscript during the editing process and Richard Lewthwaite for proof reading the English manuscript. This study was funded by the Kadoorie Farm and Botanical Garden and the Hong Kong Special Administrative Region, China.
Blake JG, Loiselle BA. 1991. Variation in resource abundance affects capture rates of birds in three lowland habitats in Costa Rica. Auk, 108: 114–130.
Blake JG, Loiselle BA. 2000. Diversity of birds along an elevational gradient in the cordillera central, Costa Rica. Auk, 117: 663–686.
Blake JG, Loiselle BA. 2001. Bird assemblages in second-growth and old-growth forests, Costa Rica: Perspectives from mist nets and point counts. Auk, 18: 304–326.
Cheng TH, Tan YK. 1973. Birds of Hainan Island Ⅱ. Acta Zool Sin, 19: 405–416. (in Chinese)
Dunn EH, Ralph CJ. 2004. Use of mist nets as a tool for bird population monitoring. Stud Avian Biol, 29: 1–6.
Fitzgerald BM, Robertson HA, Whitaker AH. 1989. Vertical distribution of birds mist-netted in mixed lowland forest in New Zealand. Notornis, 36: 311–321.
Gotelli NJ, Entsminger GL. 2001. EcoSim: Null models software for ecology. 7th ed. Acquired Intelligence Inc. and Kesey-Bear. . Accessed 20 February 2009.
Jenni L, Leuenberger M, Rampazzi F. 1996. Capture efficiency of mist nets with comments on their role in the assessment of passerine habitat use. J Field Ornithol, 67: 263–274.
Karr JR. 1971. Structure of avian communities in selected Panama and Illinois habitats. Ecol Monogr, 41: 207–233.
Karr JR. 1981. Surveying birds with mist nets. Stud Avian Biol, 6: 62–67.
Karr JR. 1990. Avian survival rates and the extinction process on Barro Colorado Island, Panama. Conserv Biol, 4: 391–397.
Lövei GL, Csörgõ T, Miklay G. 2001. Capture efficiency of small birds by mist nets. Ornis Hungarica, 11: 19–25.
Pagen RW, Thompson Ⅲ FR, Burhans DE. 2002. A comparison of point-count and mist-net detections of songbirds by habitat and time-of-season. J Field Ornithol, 273: 53–59.
Petit DR, Petit LJ, Smith KG. 1992. Habitat associations of migratory birds overwintering in Belize, Central America. In: Hagan Ⅲ JM, Johnston DW (eds) Ecology and Conservation of Neotropical Migrant Landbirds. Smithsonian Institution Press, Washington D.C., pp 247–256.
Piratelli A. 2003. Mesh size and bird capture rates in mato grosso do sul state, Brazil Braz J Biol, 63: 105–111.
Ralph CJ, Geupel GR, Pyle P, Martin TE, DeSante DF. 1993. Handbook of field methods for monitoring landbirds. General Technical Report PSW-GTR-144, Pacific Southwest Research Station, Albany, CA.
Rappole JH, Winker K, Powell GVN. 1998. Migratory birds habitat use in southern Mexico: mist nets versus point counts. J Field Ornithol, 69: 635–643.
Remsen JV, Good DA. 1996. Misuse of data from mist-net captures to assess relative abundance in bird populations. Auk, 113: 381–398.
Terborgh J, Robinson SK, Parker Ⅲ TA, Munn CA, Pierpont N. 1990. Structure and organization of an Amazonian forest bird community. Ecol Monogr, 60: 213–238.
Wang Y, Finch D. 2002. Consistency of mist netting and point counts in assessing landbird species richness and relative abundance during migration. Condor, 104: 59–72.
Whitman AA, Hagan Ⅲ JM, Brokaw NVL. 1997. A comparison of two bird survey techniques used in a subtropical forest. Condor, 99: 955–965.
Xu LH, Liu ZH, Liao WP, Li XH, Yu SM, Qiu JC, Zhou YY, Guang GX, Lu JZ, Yan K. 1983. Birds and Beasts of Hainan Island. Science Press, Beijing. (in Chinese)
Zeng QB, Li YD, Chen BF, Wu ZM, Chen BF. 1995. A list of bio-species in Jianfengling of China. The Chinese Academy of Forestry, International Tropical Organization and The Forestry Bureau of Hainan Province. China Forestry Publishing House, Beijing. (in Chinese)
Zeng QB, Li YD, Chen BF, Wu ZM, Zhou GY. 1997. Tropical Forest Ecosystems Research and Management. China Forestry Publishing House, Beijing. (in Chinese)
Zheng GM. 1995. Ornithology. Beijing Normal University Press, Beijing. (in Chinese)
Zheng GM. 2005. A Checklist on the Classification and Distribution of the Birds of China. Science Press, Beijing. (in Chinese)
Table
2.
Species list of birds captured at Jianfengling a
Common name
Scientific name
Captures b
NV c
NT d
M e
Activity height (m) f
Emerald Dove
Chalcophaps indica
4
−
−
R
−
Cuckoo Owl
Glaucidium cuculoides
8
4
20
R
10.7 ± 3.0 (3)
Collared Pygmy Owl
Glaucidium brodiei
1
4
17
R
11.7 ± 3.3 (3)
Red-headed Trogon
Harpactes erythrocephalus
2
25
45
R
9.9 ± 1.2 (17)
Blue-bearded Bee-eater
Nyctyornis athertoni
1
16
47
R
14.9 ± 4.9 (7)
Black-browed Barbet
Megalaima oorti
2
13
158
R
13.5 ± 2.4 (10)
Silver-breasted Broadbill
Serilophus lunatus
26
66
68
R
12.0 ± 1.4 (12)
Blue-rumped Pitta
Pitta soror
3
3
3
R
0 (2)
White-throated Bulbul
Alophoixus pallidus
46
265
480
R
8.3 ± 1.9 (105)
Green-winged Bulbul
Hypsipetes mcclellandii
5
28
52
R
12.2 ± 1. 8 (13)
Chestnut Bulbul
Hemixos castanonotus
5
123
354
R
11.3 ± 3.0 (27)
Black Drongo
Dicrurus macrocercus
1
15
19
R
16.0 ± 1.0 (8)
Bronzed Drongo
Dicrurus aeneus
5
95
139
R
16.6 ± 4.2 (41)
Greater Racket-tailed Drongo
Dicrurus paradiseus
2
37
43
R
12.6 ± 2.1 (15)
Yellow-breasted Green Magpie
Cissa hypoleuca
2
−
−
R
−
Gray Treepie
Dendrocitta formosae
1
9
30
R
13.0 (5)
Racket-tailed Treepie
Temnurus temnurus
1
12
13
R
14.9 ± 2.4 (7)
Red-tailed Robin
Luscinia sibilans
8
1
1
W
0.5 (1)
Red-flanked Bush Robin
Luscinia cyanurus
9
2
2
W
5.0 (1)
White-rumped Shama
Copsychus malabaricus
11
16
19
R
3.9 ± 1.3 (6)
White-crowned Forktail
Enicurus leschenaulti
13
30
86
R
0 (22)
Orange-headed Ground Thrush
Zoothera citrina
11
−
−
R
−
Golden Mountain Thrush
Zoothera dauma
2
1
1
W
0 (1)
Grey Thrush
Turdus cardis
12
4
4
W
0 (1)
Blue-and-white Flycatcher
Cyanoptila cyanomelaena
1
1
1
W
−
Fujian Niltava
Niltava davidi
1
1
1
W
−
Hainan Blue Flycatcher
Cyornis hainanus
13
37
81
R
4.0 ± 0.5 (26)
White-throated Fantail
Rhipidura albicollis
4
19
24
R
4.5 ± 0.8 (14)
Black-naped Monarch
Hypothymis azurea
13
32
33
R
8.8 ± 1.3 (21)
Lesser Necklaced Laughingthrush
Garrulax monileger
4
5
6
R
6.5 ± 5.0 (2)
Greater Necklaced Laughingthrush
Garrulax pectoralis
3
9
22
R
8.1 ± 2.1 (5)
Grey Laughingthrush
Garrulax maesi
1
23
129
R
3.7 ± 1.7 (3)
Black-throated Laughingthrush
Garrulax chinensis
8
42
197
R
3.0 ± 0.9 (15)
Large Scimitar Babbler
Pomatorhinus hypoleucos
1
2
9
R
0.3 (1)
Rufous-necked Scimitar Babbler
Pomatorhinus ruficollis
42
46
217
R
1.1 ± 0.3 (18)
Lesser Wren Babbler
Napothera epilepidota
3
2
2
R
1.2 (1)
Rufous-capped Babbler
Stachyris ruficeps
17
79
142
R
4.9 ± 1.2 (19)
Spot-necked Babbler
Stachyris striolata
7
2
2
R
0.5 (1)
Gould's Fulvetta
Alcippe brunnea
23
73
258
R
2.1 ± 0.4 (28)
Grey-cheeked Fulvetta
Alcippe morrisonia
227
1376
1687
R
3.5 ± 0.2 (178)
White-bellied Yuhina
Erpornis zantholeuca
24
459
553
R
7.5 ± 0.6 (74)
Grey-headed Parrotbill
Paradoxornis gularis
10
112
112
R
10.3 ± 2.5 (6)
Dusky Warbler
Phylloscopus fuscatus
1
2
2
W
6.0 (1)
Pale-legged Leaf Warbler
Phylloscopus tenellipes
2
3
3
P
3.5 ± 1.5 (2)
Blyth's Leaf Warbler
Phylloscopus reguloides
1
−
−
W
−
a Classification based on Zheng (2005). b Captures: capture number. c NV: Visual observation number. d NT: Total observation number (visual + aural). e Migratory status based on Xu et al. (1983): resident (R), winter visitor (W) and passage migrant (P). f Sample number in brackets.

 DownLoad:
DownLoad:





 Email Alerts
Email Alerts RSS Feeds
RSS Feeds