
Figure
1.
Location of the Hara Biosphere Reserve in southern Iran
| Citation: |
Manman CAO, Naifa LIU, Xiaoli WANG, Mengmeng GUAN. 2010: Genetic diversity and genetic structure of the Daurian Partridge (Perdix dauuricae) in China, assessed by microsatellite variation. Avian Research, 1(1): 51-64. DOI: 10.5122/cbirds.2009.0005

|
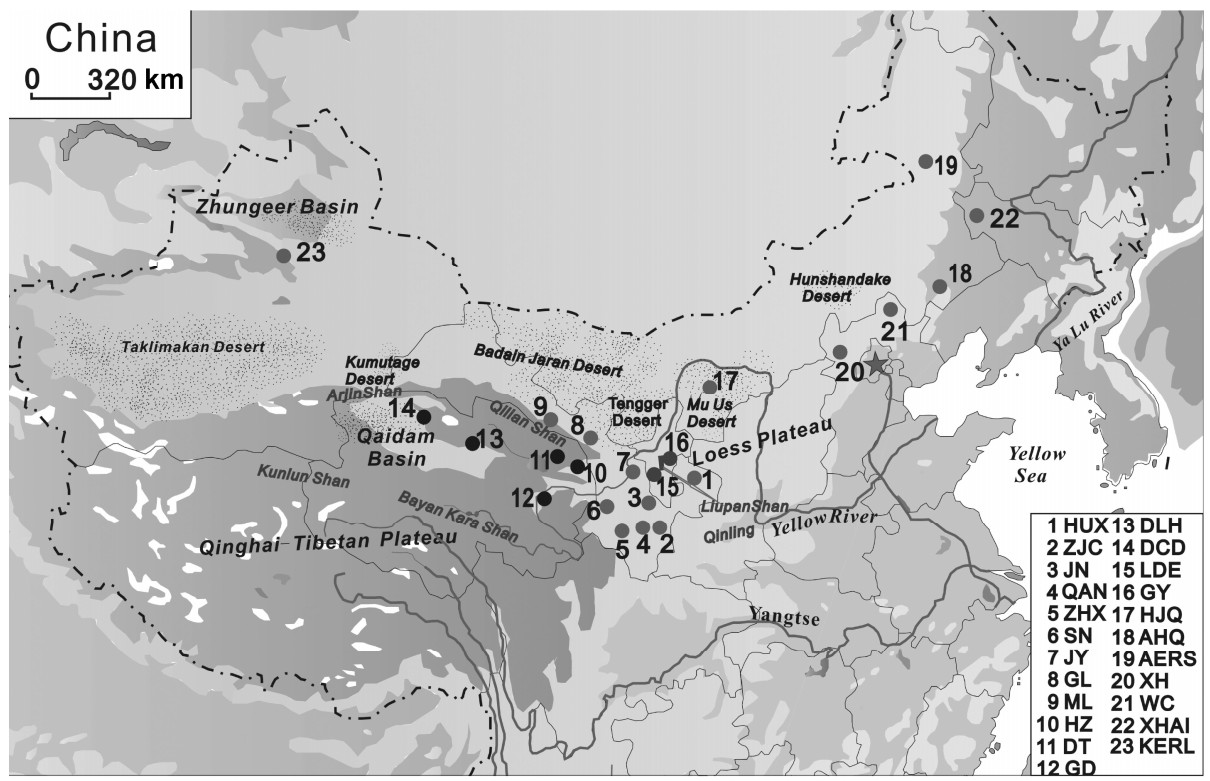
The Daurian Partridge (Perdix dauuricae) is a kind of hunting bird with high economic value. Genetic diversity and structure in the Daurian Partridge were studied by analyzing eight microsatellite loci in 23 populations found throughout the range of the species in China. The objectives were to evaluate the consequences on genetic diversity and differentiation of Daurian Partridge populations and to obtain a profound genetic insight for future management decisions and for effective measures to protect and exploit Daurian Partridges. The results showed that microsatellites were polymorphic in all Daurian Partridge populations, with a high level of genetic diversity over all the loci, especially in the Qaidam Basin populations which have the highest level of diversity. Significant genetic divergence was observed among different groups as well as between populations within the same group; most pairwise FST values were highly significant. Both phylogenetic trees and Bayesian clustering analyses revealed clear differentiation among the 23 populations of the Daurian Partridge, which were classified into two genetically differentiated groups. A bottleneck analysis indicated that Daurian Partridge populations have experienced a recent bottleneck. Our study argues that the Qaidam populations, North China populations, JN population, ZJC population, and Liupan Mountain populations should be paid special attention in order to retain adequate population sizes for maintaining genetic diversity.
Pollution of the natural environment by heavy metals is a worldwide problem. Heavy metals enter the aquatic ecosystem through a variety of anthropogenic sources as well as from natural processes (Ebrahimpour and Mushrifah, 2010). They are a serious threat because of their toxicity, bioaccumulation, long persistence and bio-magnification in the food chain (Erdoĝrul and Ates, 2006). The degree of toxic metal uptake, translocation and eventual detoxification within an organism depends on metal speciation, but also differs strongly among organisms (Mukherjee and Nuorteva, 1994; Doyle and Otte, 1997). Assessing ecosystem health adequately by means of biomonitoring requires the selection of representative indicator species. Birds are widely used to biomonitoring variation in environmental levels of anthropogenic pollutants (Furness and Camphuysen, 1997; Barbieri et al., 2009), because they are exposed to a wide range of chemicals and occupy high trophic levels and can therefore provide information on the extent of contamination in the entire food chain (Furness and Camphuysen, 1997; Burger et al., 2007).
According to Burger and Gochfeld(2000a, 2000b) feathers are useful for measuring heavy metal contamination in birds because birds sequester heavy metals in their feathers, where the proportion of the body burden is relatively constant for each metal. In general, metals in breast feathers are representative of circulating concentrations in the blood stream only during the limited time period of feather formation, which in turn represents both local exposure and mobilization from internal tissues (Lewis and Furness, 1991; Monteiro, 1996). Many studies have recommended herons and egrets as bioindicators for heavy metals in aquatic systems and local pollution around breeding sites (Boncompagni et al., 2003; Kim and Koo, 2007). Herons and gulls are high at the top of their food pyramid and can yield information over a large area around each sampling site, not only on bioavailability of contaminants but also on how, where, and when they are transferred within the food web (Battaglia et al., 2005). Hence, the aim of the present study was to investigate the level of nickel in feathers of two bird species, the Western Reef Heron (Egretta gularis) and the Siberian Gull (Larus heuglini), in order 1) to compare metal concentrations between two species with their life strategy and 2) to examine the species and gender related variation in trace nickel accumulations in the Hara Biosphere Reserve of southern Iran.
The Hara Biosphere Reserve (26°40′–27°N, 55°21′–55°52′E) is located in southern Iran, in the Straits of Khuran between Queshm Island and the Persian Gulf (Fig. 1). This area became part of the Man and Biosphere Program (MAB) of UNESCO in 1977 (UNESCO, 2010). As well, this region is one of the protected areas in Iran introduced by the Department of the Environment. The entire region was selected as wetland of international importance under the Wetlands of International Importance category as a Habitat of Aquatic Birds. This region was also introduced as one of the important bird areas by the International Organization of Birdlife (Neinavaz et al., 2010).
During November and December 2010, under license of the Environmental Protection Agency of Hormozgan, a total of thirty birds were shot and removed from throughout the Hara biosphere reserve. The collection included the Western Reef Heron (Egretta gularis) (n = 15) and the Siberian Gull (Larus heuglini) (n = 15). The birds were transported to the laboratory packed in ice. The specimen were killed, weighed, stored in plastic bags and kept at −20℃ until dissection and analysis. We chose breast feathers because they are representatives of the plumage and are less affected by molt compared to flight feathers. All feathers were analyzed in the Laboratory of the Inland Water Aquaculture Research Institute in port Anzali. The feather samples were digested in a nitric acid (HNO3) and perchloric acid (HClO4) mixture. Feathers were then accurately weighed in 150-mL Erlenmeyer flasks, where 10 mL nitric acid (65%) was added to each sample. The samples were left overnight to be slowly digested; thereafter, 5 mL perchloric acid (70%) was added to each sample. Digestion was performed on a hot plate (sand bath) at 200℃. After that, the digested samples were diluted by 25 mL deionized water. The concentration of nickel was estimated using a Shimadzu AA 680 flame atomic absorption spectrophotometer. The accuracy of the analysis was checked by measuring CRM certified reference tissue (DORM-2, NRC Canada). The detection limit for nickel was 0.039 µg·g−1. The results for nickel gave a mean recovery of 98.6%.
A statistical analysis was carried out using SPSS (version 18.0). We used a three-way ANOVA for nickel (sex, age, species, interaction (sex × age × species)). Data were log transformed to obtain normal distributions that satisfied the homogeneity of variance required by ANOVA (Custer et al., 2003; Kim et al., 2009). The nickel concentration in feathers was tested for mean differences between species using Student t tests. The level of significance was set at α = 0.05. The concentration of nickel in feathers was expressed in microgram per gram of dry weight (dw). Values are given in means ± standard errors (SE).
Variations in the nickel concentration of feathers of the Western Reef Heron and Siberian Gull are presented in Table 1. The results show that there was a significant difference between the mean nickel concentrations in the two bird species, i.e. the Western Reef Heron and Siberian Gull, while there was no evidence of significant different accumulation between genders and ages (Table 2). Also, the results indicate that the level of nickel concentration in the Western Reef Heron was higher in females than in males; in contrast, the level of nickel concentration in the Siberian gull was higher in males.
| Species | Male/adult | Male/juvenile | Female/adult | Female/juvenile |
| Western Reef Heron | ||||
| Geometric mean | 2.77 | – | 3.79 | 4.67 |
| Mean ± SE | 3.47±2.7 | – | 4.67±2.7 | 4.67 |
| Number | 9/9 | – | 5/5 | 1 |
| Siberian Gull | ||||
| Geometric mean | 8.89 | 3.87 | 6.72 | 7.24 |
| Mean ± SE | 8.94±1.1 | 4.38±2.5 | 7.6±3.4 | 8.1±4.8 |
| Number | 3/3 | 3/3 | 6/6 | 3/3 |
 DownLoad:
CSV
DownLoad:
CSV
| Source of variation | Mean square | F | p |
| Species | 78.23 | 8.56 | 0.01 a |
| Gender | 9.56 | 1.04 | NS b |
| Age | 7.57 | 0.82 | NS |
| Intercept (species × gender × age) | 572.55 | 62.66 | 0.001 |
|
a p-value for 3-way ANOVA. Interaction term significant as indicated. b NS = not significant at p > 0.05. |
|||
DownLoad:
CSV
Nickel is not an important trace element in organisms, but at high levels, they can cause adverse health effects. Sources of heavy metals vary considerably. It is emitted into the environment by both natural and man-made sources. Once released into the environment, nickel readily forms complexes with many ligands, making it more mobile than most heavy metals (Mansouri et al., 2011). Nickel is related to the pigmentation of feathers in birds and excreted via the feathers by moulting (Honda et al., 1986). Nickel concentrations in the current study (3.47–8.94 µg·g−1) were higher than those in Fulica atra (0.8 µg·g−1), Phalacrocorax carbo (0.5 µg·g−1) and Nycticorax nycticorax (1.2 µg·g−1) from Russia (Lebedeva, 1997), in Egretta alba (0.2 µg·g−1) from Korea (Honda et al., 1986) and Parus major (0.25 µg·g−1) from China (Deng et al., 2007). In our study, the nickel concentrations were similar to those in Bubulcus ibis L. (7.8–9.0 µg·g−1) from Pakistan (Malik and Zeb, 2009).
Few studies have examined the effect of gender on the accumulation of heavy metals in feathers and other tissues (Burger, 1995; Zamani-Ahmadmahmoodi et al., 2010). Several studies reported no significant differences in the heavy metal content of feathers between male and female birds (Hutton, 1981; Zamani-Ahmadmahmoodi et al., 2009a). Similarly, in the present study there was no evidence of significant different accumulations between male and female birds (Table 2), suggesting that both sexes utilize similar foraging strategies in both species (Hindell et al., 1999). While studying heavy metals in the feathers of Larus dominicanus, Barbieri et al. (2009) showed that the levels of nickel concentration were higher in adults (5.92 µg·g−1) than in juveniles (2.23 µg·g−1). Similar levels of nickel have been detected in other seabird species from different parts of the world (Norheim, 1987). Adults have had several years to accumulate metals in their internal tissues; these can be mobilized into the blood and deposited in feathers during their formation (Burger, 1994). Elsewhere, Burger and Gochfeld (1991) pointed to heavy metal concentrations in feathers of adult birds that may reflect exposure obtained at other time of the year, including exposure at non-breeding areas. On the other hand, while they were studying heavy metal concentrations in the feathers of Herring Gulls (Larus argentatus) in Captree, Long Island, they showed that the cadmium concentration was higher in juveniles but the lead concentration was higher in adults (Burger, 1995). Differences in levels of metal concentrations in adults and fledglings might also occur if adults and young eat different types of food during the breeding season, or different-sized food items (Burger, 1996).
Research has indicated that the concentration of heavy metals in the tissues of migratory birds is higher than that in resident birds (Pacyna et al., 2006; Zamani-Ahmadmahmoodi et al., 2009b). Siberian Gulls are winter visitors to Hara Biosphere Reserve, while Western Reef Heron are residents there. The results of the current study show that the amount of nickel in the Siberian Gull feathers is higher than in the Western Reef Heron.
The results of three-way ANOVA showed there were significant differences between the Western Reef Heron and Siberian Gull (p < 0.01). The Siberian Gull showed higher nickel concentrations than the Western Reef Heron. Birds that are large fish eaters should accumulate higher levels than those that eat a range of different foods or smaller fish. Furthermore, levels of metal in birds should reflect the levels in the fish they consume (Burger, 2002). The main difference in nickel levels between Western Reef Heron and Siberian Gull could be the result of different phylogenetic origin and physiology (Teal, 1969; Welty, 1975; Deng et al. 2007), or because they grow their feathers in different breeding areas with different levels of background contamination. Also, metabolic rates vary inversely with body weight and directly with activities such as flight and rest. Being smaller than Western Reef Heron, Siberian Gull was expected to have a higher metabolic rate. Higher metabolic rates may cause fast accumulations of trace nickel in the Siberian Gull. In general, the Siberian Gull eats more invertebrates than the Western Reef Heron, catches some larger fish (between 20–25 cm in size), consumes offal discarded by fishing boats and eats dead fish found along the shore, while the Western Reef Heron eats smaller fish, amphibians and insects.
|
Avise JC. 1994. Molecular Markers, Natural History and Evolution. Chapman & Hall, New York
|
|
Belkhir K, Borsa P, Chikhi L, Raufaste N, Catch F. 2004. GENETIX (version 4.02), Software under WindowsTM for the genetics of the populations Laboratory Genome, Populations, Interactions, CNRS UMR 5000, University of Montpellier Ⅱ, Montpellier, France
|
|
Chen Q, Chang C, Liu NF. 1999. Mitochondrial DNA introgression between two parapatric species of Alectoris. Acta Zool Sin, 45(4): 456–463 (in Chinese)
|
|
Excoffier L, Laval G, Schneider S. 2005. ARLEQUIN, version3.0: an integrated software package for population genetics data analysis. Evol Bioinfor Online, 1: 47–50
|
|
Frankel OH, Soule ME. 1981. Conservation and Evolution. Cambridge University Press, Cambridge, England
|
|
Goudet J. 2001. FSTAT. A program to estimate and test gene diversities and fixation indices, (Version 2.9.3). Updated from Goudet (1995)
|
|
Hamrick JL, Godt MJW. 1989. Allozyme diversity in plant species. In: Brown AHD, Clegg MT, Kahler AL, Weir BS (eds) Plant Population Genetics, Breeding, and Genetic Resources. Sinauer, Sunderland, MA, pp 43–63
|
|
Hartl DL, Clark AG. 1997. Principles of Population Genetics. Sinauer, Sunderland, MA
|
|
Heuertz M, Hausman JF, Hardy OJ, Vendramin GG, Frascaria-Lacoste N, Vekemans X. 2004. Nuclear microsatellites reveal contrasting patterns of genetic structure between western and southeastern European populations of the common ash (Fraxinus excelsior L. ). Evolution, 58: 976–988
|
|
Hou P, Wei M, Zhang LX, Liu NF. 2002. Genetic structure of edge population in Przewalski's rock partridge (Alectoris magna). Acta Zool Sin, 48(3): 333–338 (in Chinese)
|
|
Johnsgard PA. 1988. The Quails, Partridges, and Francolins of the World. Oxford University Press, Oxford
|
|
Liu NF, Huang ZH, Wen LY. 2004. Subspecies divergence of Przewalski's rock partridge (Alectoris magna) description of a new subspecies. Acta Zootaxon Sin, 29(3): 600–605 (in Chinese)
|
|
Liu NF, Wang XT, Luo WY, Chang C. 1988. Avifauna study of the Liupan Mountain in Ningxia. J Lanzhou Univ Nat Sci Ed, 24: 63–75 (in Chinese)
|
|
Luikart G, Cornuet JM. 1997. Empirical evaluation of a test for identifying recently bottlenecked populations from allele frequency data. Conserv Biol, 12: 228–237
|
|
Madge S, McGowan P. 2002. Pheasants, Partridges and Grouse: A Guide to the Pheasants, Partridges, Quails, Grouse, Guineafowl, Buttonquails and Sandgrouse of the World. Christopher Helm, London
|
|
Mantel N. 1967. The detection of disease clustering and a generalized regression approach. Cancer Res, 27: 209–220
|
|
Nei M. 1987. Molecular Evolutionary Genetics. Columbia University Press, New York
|
|
Sun HY, Cao ZW, Jiang HL, Yu LG. 1998. Brooding and growth of the younger Perdix dauuricae. Forest Sci Technol, 23(4): 28–30 (in Chinese)
|
|
Sun HY, Ge DN, Hong YT, Cao ZW. 1996. Research about domesticating and breeding technology of Daurian Partridge (Perdix dauuricae). Forest Sci Technol, 21(4): 32–35 (in Chinese)
|
|
Turner PC, McLennan AG, Bates AD, White MRH. 2003. Instant Notes in Molecular Biology, 2nd edn. Science Press, Beijing
|
|
Weir BS, Cockerham CC. 1984. Estimating F-statistics for the analysis of population structure. Evolution, 38: 1358–1370
|
|
Woodruff DS. 1989. The problems of conserving genes and species. In: Weston D, Pearl M (eds) Conservation for the Twenty First Century. Oxford University Press, New York, pp 76–88
|
|
Zhang ZW, Liang W, Sheng G. 1994. Studies on the nest site selection of Daurian partridge. Zool Res, 15(4): 37–43 (in Chinese)
|
|
Zhang ZW, Wu YC. 1992. The Daurian Partridge (Perdix dauuricae) in north-central China. Gibier Faune Sauvage, 9: 591–595
|
|
Zhang ZW, Liang W. 1997. Breeding ecology of Daurian Partridge (Perdix dauuricae) in Shanxi. Chinese J Zool, 2(2): 23–25 (in Chinese)
|
|
Zhao ZJ, Zhang S, Feng KF. 1992. The biology of the Daurian Partridge (Perdix dauuricae suschkini) in northeastern China. Gibier Faune Sauvage, 9: 597–604
|
|
Zheng ZX. 1978. Fauna Sinica. Aves. Galliformes. Science Press, Beijing (in Chinese)
|
| Xiaocai Tan, Shilong Liu, Eben Goodale, Aiwu Jiang. 2022: Does bird photography affect nest predation and feeding frequency?. Avian Research, 13(1): 100036. DOI: 10.1016/j.avrs.2022.100036 | |
| Limin Wang, Ghulam Nabi, Liyun Yin, Yanqin Wang, Shuxin Li, Zhuang Hao, Dongming Li. 2021: Birds and plastic pollution: recent advances. Avian Research, 12(1): 59. DOI: 10.1186/s40657-021-00293-2 | |
| Juan Arizaga, Daniel Alonso, Ariñe Crespo, Xabier Esparza, Efrén Fernández, Iñigo López, David Martín, Antonio Vilches. 2020: Yearly variation in the structure and diversity of a non-breeding passerine bird community in a Mediterranean wetland. Avian Research, 11(1): 29. DOI: 10.1186/s40657-020-00215-8 | |
| Yan Zhou, Anping Chen, Jenny Q. Ouyang, Yanlin Liu, Aiyu Zheng, Zaixi Yang, Yong Zhang, Bin Wang, Yifei Jia, Shengwu Jiao, Qing Zeng, Changhu Lu. 2020: Comparing community birdwatching and professional bird monitoring with implications for avian diversity research: a case study of Suzhou, China. Avian Research, 11(1): 19. DOI: 10.1186/s40657-020-00205-w | |
| Ali M. Zanaty, Ahmed M. Erfan, Wessam H. Mady, Fatma Amer, Ahmed A. Nour, Neveen Rabie, Mohamed Samy, Abdullah A. Selim, Wafaa M. M. Hassan, Mahmoud M. Naguib. 2019: Avian influenza virus surveillance in migratory birds in Egypt revealed a novel reassortant H6N2 subtype. Avian Research, 10(1): 41. DOI: 10.1186/s40657-019-0180-7 | |
| Lidan Zhao, Lijun Gao, Wenyu Yang, Xianglong Xu, Weiwei Wang, Wei Liang, Shuping Zhang. 2017: Do migrant and resident species differ in the timing of increases in reproductive and thyroid hormone secretion and body mass?A case study in the comparison of pre-breeding physiological rhythms in the Eurasian Skylark and Asian Short-toed Lark. Avian Research, 8(1): 10. DOI: 10.1186/s40657-017-0068-3 | |
| Anders Pape Møller. 2015: The allometry of number of feathers in birds changes seasonally. Avian Research, 6(1): 2. DOI: 10.1186/s40657-015-0012-3 | |
| Jörg HOFFMANN, Udo WITTCHEN, Ulrich STACHOW, Gert BERGER. 2013: Identification of habitat requirements of farmland birds based on a hierarchical structured monitoring scheme. Avian Research, 4(4): 265-280. DOI: 10.5122/cbirds.2013.0026 | |
| Shuihua CHEN, Qin HUANG, Zhongyong FAN, Cangsong CHEN, Yiwei LU. 2012: The update of Zhejiang bird checklist. Avian Research, 3(2): 118-136. DOI: 10.5122/cbirds.2012.0017 | |
| Dongsheng LI, Corwin SULLⅣAN, Zhonghe ZHOU, Fucheng ZHANG. 2010: Basal birds from China: a brief review. Avian Research, 1(2): 83-96. DOI: 10.5122/cbirds.2010.0002 |
Figures(3) / Tables(7)





| Species | Male/adult | Male/juvenile | Female/adult | Female/juvenile |
| Western Reef Heron | ||||
| Geometric mean | 2.77 | – | 3.79 | 4.67 |
| Mean ± SE | 3.47±2.7 | – | 4.67±2.7 | 4.67 |
| Number | 9/9 | – | 5/5 | 1 |
| Siberian Gull | ||||
| Geometric mean | 8.89 | 3.87 | 6.72 | 7.24 |
| Mean ± SE | 8.94±1.1 | 4.38±2.5 | 7.6±3.4 | 8.1±4.8 |
| Number | 3/3 | 3/3 | 6/6 | 3/3 |
DownLoad:
CSV
| Source of variation | Mean square | F | p |
| Species | 78.23 | 8.56 | 0.01 a |
| Gender | 9.56 | 1.04 | NS b |
| Age | 7.57 | 0.82 | NS |
| Intercept (species × gender × age) | 572.55 | 62.66 | 0.001 |
|
a p-value for 3-way ANOVA. Interaction term significant as indicated. b NS = not significant at p > 0.05. |
|||
DownLoad:
CSV
 Email Alerts
Email Alerts RSS Feeds
RSS Feeds